Свернутая полипептидная цепь
Cтраница 1
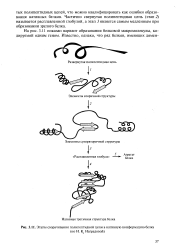 |
Этапы сворачивания полипептидной цепи в нативную конформацию белка. [1] |
Частично свернутая полипептидная цепь ( этап 2) называется расплавленной глобулой, а этап 3 является самым медленным при образовании зрелого белка. [2]

Третичная структура свернутой полипептидной цепи стабилизируется целым рядом нековалентных взаимодействий ( особенно гидрофобными взаимодействиями между неполярными R-группами), электростатическим притяжением между противоположно заряженными R-группами и водородными связями. Все эти взаимодействия, будучи слабыми по своей природе, в совокупности оказываются очень прочными. В некоторых глобулярных белках определенную роль в формировании и стабилизации третичной структуры играют дисуль-фидные поперечные связи. Информация, определяющая третичную структуру белков, заложена в аминокислотной последовательности их полипептидных цепей, о чем свидетельствует тот факт, что гомологичные белки из разных видов имеют не только много общих инвариантных аминокислотных остатков, но и одинаковую конформацию. Доказательством этого утверждения служит то, что многие денатурированные глобулярные белки, утратившие свойственную им биологическую активность, способны самопроизвольно ренатурировать с полным восстановлением их биологической активности. [4]
В растворе молекула муцина представляет собой довольно жесткий клубок, состоящий из гибкой, беспорядочно свернутой полипептидной цепи с заключенными внутрь молекулами воды. [5]
В класс II попали белки со значением), приблизительно равным его среднему значению для беспорядочно свернутых полипептидных цепей, что, вероятно, свидетельствует о неупорядоченности их конфор-мации. Наконец, в класс III были включены белки со значением АО ниже среднего значения для беспорядочно свернутых полипептидных цепей - можно было предполагать, что это связано с наличием ( 3-структуры или левых а-спиралей, а быть может, и элементов каких-то неизвестных структур. [6]
Денатурация, по-видимому, характеризуется некоторым общим расслаблением довольно жестко закрепленных структурных отношений, характеризующих нативный белок. Это приводит к раскручиванию свернутых полипептидных цепей ( стр. [7]
В молекуле гемоглобина гем-группы расположены далеко друг от друга ( 3000 пм), поэтому прямого взаимодействия между ними недостаточно, чтобы объяснить это явление. Однако было найдено, что присоединение кислорода или другого лиганда изменяет форму свернутой полипептидной цепи и благодаря взаимодействию между соседними цепями этот эффект передается. [8]
В класс II попали белки со значением), приблизительно равным его среднему значению для беспорядочно свернутых полипептидных цепей, что, вероятно, свидетельствует о неупорядоченности их конфор-мации. Наконец, в класс III были включены белки со значением АО ниже среднего значения для беспорядочно свернутых полипептидных цепей - можно было предполагать, что это связано с наличием ( 3-структуры или левых а-спиралей, а быть может, и элементов каких-то неизвестных структур. [9]
Если температура может влиять на высшие порядки структуры белков, то мы можем предсказать, что термолабильной окажется и конформация растущей ( синтезируемой) полипептидной цепи, связанной с комплексом полирибосомы. Более того, растущий полипептид, который еще не приобрел своей окончательной конформации, может быть даже более чувствителен к температуре, чем полностью свернутая полипептидная цепь. [10]
Спираль и р-конформация возникают самопроизвольно и остаются устойчивыми лишь в том случае, если набор расположенных по соседству аминокислотных остатков в полипептидной цепи имеет соответствующую последовательность R-групп. Третичная структура глобулярных белков тоже обусловлена. Образование изгибов полипептидной цепи, а также направление и угол поворота цепи в этих изгибах обусловлены числом и положением определенных аминокислотных остатков, таких, как пролин, треонин и серии, которые способствуют образованию изгибов. Более того, как мы увидим дальше, петли плотно свернутой полипептидной цепи сохраняют характерное для них положение в пространстве также благодаря разного рода взаимодействиям между R-группами соседних петель. [11]
Спираль имеет правое вращение, при этом объемные боковые цепи ь-аминокислот направлены от центра спирали. В ( 5-структуре полиамидные цепи располагаются рядом в антипараллельном положении, образуя слой полипептидных цепей, связанных между собой водородными связями. Участки - структуры могут образовываться и внутри одной белковой молекулы за счет складывания свернутой полипептидной цепи. [12]
Белки-это самый многочисленный класс присутствующих в клетках макромолекул; они составляют свыше половины сухого веса клеток. Белки представляют собой очень длинные полипептидные цепи, содержащие от 100 до 1 000 и более аминокислотных звеньев, соединенных друг с другом пептидными связями. Простые белки в процессе гидролиза распадаются только на аминокислоты; сложные белки содержат еще и другие дополнительные компоненты-ионы металлов или органические простетические группы. Одни белки имеют волокнистую ( фибриллярную) структуру и нерастворимы, тогда как другие состоят из плотно свернутых полипептидных цепей и имеют глобулярную форму. [13]
Белковая структура гемоглобина более сложна. Гемоглобин млекопитающих ( в том числе человека) имеет мол. Преобладающая форма гемоглобина у взрослых людей - гемоглобин А - имеет две пары полипептидных цепей: а-цепей, каждая из которых состоит из 141 аминокислотного остатка, и р-цепей - по 146 остатков в каждой. В содержащемся в небольшом количестве у взрослых людей гемоглобине А % и в зародышевом гемоглобине F Р - цепи замещены другими полипептидами. Несмотря иа то что аминокислотные последовательности полипептидных цепей гемоглобина и миоглобина в значительной степени различаются, трехмерные структуры их чрезвычайно сходны и темы в молекулах того и другого занимают гидрофобные полости внутри свернутых полипептидных цепей. Проксимальный и дистальный остатки гистидина входят в число девяти аминокислот, которые одинаковы во всех миоглобинах и гемоглобинах у ряда изученных в этом отношении видов животных. [14]
С; сохраняются длит, время только в высушенном виде. При единичном ужале-нии обычно выделяется менее 1 мг яда. Яды содержат полипептиды, ферменты ( фосфолипазы А и В, кислая фосфатаза, фосфодиэстераза, ацетилхолинэстераза, 5 -нуклеотидаза, гиалуронидаза, рибонуклеаза), моно - и полисахариды; в нек-рых ядах найдены серотонин и гистамин. Уникальное св-во ядов нек-рых видов ( напр. ВШШае и Зсог - рюгшЗае) - наличие нейротоксинов, избирательно действующих на млекопитающих, насекомых или ракообразных. Токсины млекопитающих обычно имеют мол. Их свернутая полипептидная цепь ( 60 - 70 аминокислотных остатков) стабилизирована неск. Нейро-токсины скорпионов в неск. Их активность обусловлена модификацией № - каналов алектровозбудимых мембран. [15]
Страницы: 1