Выход - биомасса
Cтраница 1
Выход биомассы зависит, таким образом, от площади коллектора солнечной энергии ( листьев), функционирующих в течение года, и числа дней в году с такими условиями освещенности, когда возможен фотосинтез с максимальной скоростью, что определяет эффективность всего процесса. Результаты определения доли солнечной радиации ( в %), доступной растениям ( фотосинтетически активной радиации, ФАР), и знание основных фотохимических и биохимических процессов и их термодинамической эффективности позволяют рассчитать вероятные предельные скорости образования органических веществ в пересчете на углеводы. [1]
Выход биомассы в этом случае значительно выше, чем при кислотном гидролизе. Полученные дрожжи имеют белый цвет, в них содержится 49 7 - 51 1 % белка. [2]
Выход биомассы в этом случае составляет 50 - 60 % от используемых РВ. [3]
Выход биомассы можно определить и в периодическом процессе, когда максимальное количество биомассы получают в стационарной фазе роста. [4]
Резко сократится выход биомассы, снизится содержание белка в пей и увеличится количество липидов. Усилится процесс поглощения клетками парафинов, увеличатся производственные потери, снизится процесс почкования и деления клеток. [5]
Условием линейности зависимости выхода биомассы от концентрации субстрата в питательной среде, согласно формуле (2.63), является постоянство как экономического коэффициента У, так и константы равновесия процесса размножения К. Это значит, что даже при yconst изменение запаса субстрата в питательной среде не всегда будет сопровождаться пропорциональным изменением равновесной концентрации микроорганизмов. [6]
В общем случае коэффициент выхода биомассы является переменной величиной, зависящей от условий культивирования. Вариабельность этого коэффициента очень важна с точки зрения оптимизации процесса. [7]
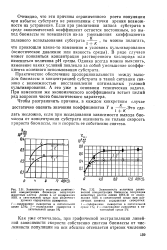
Практическое обеспечение пропорциональности между выходом биомассы и концентрацией субстрата в такой ситуации связано с возможностью восстановления оптимальных условий культивирования. [9]
С ослаблением активности культуры снижается выход биомассы из единицы питательных веществ. [10]
Это должно сопровождаться соответствующим повышением выхода биомассы, пока какие-либо новые лимитирующие факторы процесса культивирования не станут значимыми на фоне высокой концентрации субстрата и не вызовут снижения коэффициента его полезного использования. [11]
Таким образом, задача повышения выхода биомассы, решение которой в первую очередь связано с балансировкой состава питательной среды и соответствующим [ согласно условию ( XXXIII) и ( XXXIV) ] увеличением в ней концентраций компонентов, во многих случаях влечет за собой необходимость создания системы дозаторов для жидких веществ и интенсификации массообмена для газообразных. [12]
В этом случае заметно уже снижение выхода биомассы при существенном увеличении значений а ыансо3 и снижении а ог - Повышение концентрации кислорода в газовой фазе ( обогащение кислородом аэрирующего воздуха) при k 0 28 ч - 1 восстанавливает выход биомассы, но характер энергетического обмена и в этих условиях остается сдвинутым в сторону гликолиза по сравнению с обменом животных клеток, культивируемых в условиях более интенсивного массообмена по кислороду. [13]
 |
Мицелиальные грибы. культуры дрожжей часто да. [14] |
Но при изучении скорости размножения и выхода биомассы указанных мицелиальных грибов на гидролизных и сульфитно-спиртовых средах при сильной и слабой аэрации А. П. Крючкова и П. Н. Фишер пришли к выводу, что мицелиальные грибы значительно медленнее накапливают биомассу, чем пентозные дрожжи. Но если, например, мицелиаль-ныи rpHg у з культивировать совместно с дрожжеподоб-ным грибком Л-2, то он хорошо растет и усваивает органические вещества барды, в том числе арабинозу. [15]
Страницы: 1 2 3 4