Ген-оператор
Cтраница 3
Синтез мРНК на структурных генах молекулы ДНК непосредственно контролируется определенным участком, называемым геном-оператором. Он служит как бы пусковым механизмом для функционирования структурных генов. Ген-оператор локализован на крайнем отрезке структурного гена или структурных генов, регулируемых им. Считывание генетического кода, т.е. формирование мРНК, начинается с промотора - участка ДНК, расположенного рядом с геном-оператором и являющегося точкой инициации для синтеза мРНК, и распространяется последовательно вдоль оператора и структурных генов. Синтезированную молекулу мРНК, кодирующую синтез нескольких разных белков, принято называть полигенным ( полицистронным) транскриптом. Координированный одним оператором одиночный ген или группа структурных генов образует оперон. [31]
 |
Схема оперона. [32] |
От гена-оператора зависит работа группы генов, программирующих синтез нескольких белков, в том числе р-галактозидазы. Если ген-оператор подавлен, репрессирован белком-репрессо-ром, то структурные гены не функционируют. Ген-оператор и подчиненные ему структурные гены образуют так называемый оперон. [33]
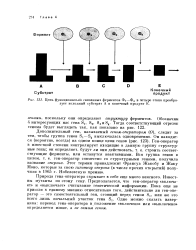 |
Цепь функционально связанных ферментов Ii - Ф4 в четыре этапа преобразует исходный субстрат А в конечный продукт Е. [34] |
Природа гена-оператора скрывает в себе еще много неясного. Известны мутанты по этому гену. Предполагается, что ген-оператор включает и выключает считывание генетической информации. Одно можно сказать наверняка: перевод гена-оператора в положение включено или выключено определяется извне, а не самим геном. [35]
Таким образом, биосинтез мРНК, контролирующий синтез белка в рибосомах, зависит от функционального состояния репрессора. Этот реп-рессор представляет собой тетрамерный белок с общей мол. Если он находится в активном состоянии, т.е. не связан с индуктором, то блокирует ген-оператор и синтеза мРНК не происходит. [36]
Пусть для этой последовательности нужно три фермента. Тогда по схеме Моно и Жакоба этот продукт вступает в реакцию с одним из белков, производимым на особом участке ДНК, называемом геном-регулятором. Это соединение, так называемый репрессорный белок, в свою очередь, действует на систему, состоящую из участка ДНК, называемого оператором ( ген-оператор), и участков, называемых структурными генами. На структурных генах и получается м - РНК, непосредственно используемая для синтеза требуемых белков-ферментов. Роль гена-оператора заключается в контролировании скорости использования структурных генов для синтеза белка. Если репрессор соединился с геном-оператором, блокировал его, то работа соответствующих структурных генов прекращается. В нашем примере ген-оператор должен контролировать деятельность трех участков ДНК, производящих нужные м - РНК и, следовательно, и ферменты для данной сложной реакции. Выключение оператора в результате фиксации на нем белка репрессора, связанного с продуктом реакции Р, прекращает я производство ферментов для реакции. Если концентрация продукта Р понизилась, оператор освободился от блокады, то синтез ферментов возобновляется и вся биохимическая машина запускается вновь. [37]
Было показано, что наряду со структурными генами, кодирующими синтез ферментов, в бактериальном геноме существуют специальные регуляторные гены. Ген-регулятор кодирует синтез специфического аллосте-рического белка-репрессора, имеющего два центра связывания: один узнает определенную последовательность нуклеотидов на участке ДНК, называемом оператором ( ген О), другой - взаимодействует с эффектором. Ген-оператор расположен рядом со структурным геном ( генами) и служит местом связывания репрессора. [38]
Отсюда вещество, которое непосредственно действует на оператор и выключает его, назвали репрессором, Репрессор состоит из вещества, продуцируемого геном-регулятором R, и низкомолекулярного вещества - в данном случае это аргинин, и по аналогии с голоферментом, состоящим из апофермента и кофермента, говорят об апорепрессоре и корепрессоре. Ген R производит апорепрессор, неэффективный сам по себе, который поступает в цитоплазму. Накопившийся корепрессор - конечный продукт цепи ферментативных реакций ( аргинин) - соединяется с апорепрессором, образуя голорепрессор. Последний же подавляет ген-оператор, а тем самым и весь оперон, контролирующий синтез аргинина. Если же, напротив, аргинин, образующийся в нашей последовательности реакций, будет немедленно перерабатываться дальше, например включаться в те или иные белки, то он не будет накапливаться и, следовательно, не сможет образовать совместно с апорепрессором активный репрессор. [39]
Индуцированный синтез подавляется специфическим веществом - репрессором, находящимся в цитоплазме. Репрессор синтезируется особым геном-регулятором. Позднее было показано, что репрессоры - белки. Репрессор действует на ген-оператор, управляющий переносом информации от нескольких структурных генов к синтезируемым белкам. При воздействии реп-рессора прекращается работа всей совокупности этих генов - всего оперона. [40]
После того, как в 1961 г. Жакоб и Моно предложили механизм, с помощью которого гены могут включаться и выключаться, были получены другие данные, позволившие прояснить различные аспекты этого механизма. На основе генетических данных было высказано предположение о существовании гена-промотора, расположенного рядом с геном-оператором и действующего между ним и геном-регулятором. Предполагается, что ген-промотор несет две функции. Это перемещение, конечно, зависит от того, находится ли ген-оператор в активном состоянии или нет. [41]
Однако в клетке имеется еще одно вещество, способное соединяться с апорепрессором; мы будем называть его индуктором. Если корепрессор присутствует в большом количестве, тогда, конечно, у индуктора нет никаких шансов соединиться с апорепрессором. Но допустим, что глюкоза расщеплена вся без остатка. Тогда концентрация корепрессора резко падает, и тут наступает черед индуктора. Если ему удается прикрепиться к апоре-прессору, то он его инактивирует. Комплекс апорепрессор-индуктор образует неактивный репрессор; теперь ген-оператор больше не может находиться в выключенном состоянии, он дерепрессируется. Со структурных генов начинает считываться информация, начинается синтез ферментов, разлагающих лактозу. Клетка приступает к потреблению лактозы. [42]
Пусть для этой последовательности нужно три фермента. Тогда по схеме Моно и Жакоба этот продукт вступает в реакцию с одним из белков, производимым на особом участке ДНК, называемом геном-регулятором. Это соединение, так называемый репрессорный белок, в свою очередь, действует на систему, состоящую из участка ДНК, называемого оператором ( ген-оператор), и участков, называемых структурными генами. На структурных генах и получается м - РНК, непосредственно используемая для синтеза требуемых белков-ферментов. Роль гена-оператора заключается в контролировании скорости использования структурных генов для синтеза белка. Если репрессор соединился с геном-оператором, блокировал его, то работа соответствующих структурных генов прекращается. В нашем примере ген-оператор должен контролировать деятельность трех участков ДНК, производящих нужные м - РНК и, следовательно, и ферменты для данной сложной реакции. Выключение оператора в результате фиксации на нем белка репрессора, связанного с продуктом реакции Р, прекращает я производство ферментов для реакции. Если концентрация продукта Р понизилась, оператор освободился от блокады, то синтез ферментов возобновляется и вся биохимическая машина запускается вновь. [43]
При распаде лактозы действуют одновременно две системы: во-первых, система разложения лактозы и, во-вторых, конститутивный путь распада глюкозы; в последний и отводятся продукты распада лактозы. Нельзя ожидать, что обе системы будут заранее в точности согласованы друг с другом. Иное дело, если будет разложено больше лактозы, чем глюкозный путь в состоянии осилить. Вот тут-то и включается тонкая настройка: галактоза - промежуточный продукт, образующийся на первом этапе разложения лактозы - накапливается все в большем количестве и начинает действовать как корепрессор. Она оттесняет индуктор ( лактозу) от апорепрессора и связывается с ним, образуя активный репрессор. В результате выключается ген-оператор, блокируется синтез ферментов и на некоторое время прекращается распад лактозы - до тех пор, пока не снизится в достаточной степени концентрация галактозы. [44]
В настоящее, время считают, что регуляции подвержены все или почти все этапы биосинтеза белков. Например, метаболиты и гормоны могут изменять сродство белков-репрессоров к регуляторным отделам ДНК; гормоны способны модифицировать активность метилаз, участвующих в биосинтезе рРНК; новообразованные белки способны активировать рибонукле-азы и тем самым ускорять распад своих мРНК и т.п. Согласно теории Жакоба и Моно, в биосинтезе белков у бактерий участвуют три типа генов: структурные гены, ген-оператор и ген-регулятор. Структурные гены определяют первичную структуру белков. Формирование мРНК начинается с промотора и далее распространяется вдоль оператора и контролируемых им структурных генов. Оператор и структурные гены называют опероном. Деятельность оперона контролируется геном-регулятором. Оперон и ген-регулятор находятся в разных участках цепи ДНК, поэтому связь между ними осуществляется с помощью белка-репрессора, синтезируемого по информации гена-регулятора. Если репрессор связан с геном-оператором, то РНК-по-лимераза не может синтезировать мРНК, а следовательно, не синтезируются и белки. Если ген-оператор свободен, процесс транскрипции возможен и информация структурных генов используется для синтеза белков. [45]
Страницы: 1 2 3 4