Суммарная каталитическая активность
Cтраница 1
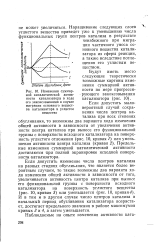 |
Изменения суммарной каталитической активности катализатора в ходе его закоксовывания в случае миграции основного вещества катализатора в углистое вещество. [1] |
Предельные изменения суммарной каталитической активности достигаются при полной экранировке исходной поверхности катализатора. [2]
Мы рассмотрим вопрос о суммарной каталитической активности в клетке и в связи с этим определим оптимальную плоидность ядра. В зависимости от того, сколько копий полипептидов разных типов имеется в клетке, будет меняться соотношение разных иэоферментов и, соответственно, суммарная ферментативная активность клетки. Здесь и далее мы будем подразумевать, что все имеющиеся в клетке полипептиды участвуют в агрегации и объединение разных типов полипептидов в белковый комплекс происходит случайно. [3]
Таким образом, наличие изоферментов позволяет варьированием соотношения аллелей изменять суммарную каталитическую активность клетки и точнее регулировать интенсивность метаболических процессов. Возможность изменения соотношения аллелей появляется только у полиплоидов, а создание растения с наибольшим приближением к максимуму активности по всем ферментам и с наиболее точной регуляцией скоростей реакций и есть создание организма с оптимальной плоид - ностью ядра. Так как большинство ферментов состоит из нескольких субъединиц, то рассмотренная связь имеет, по всей видимости, большое значение. [4]
Данные об участии парамагнитных центров молекулярных сит, насыщенных серой, в каталитических процессах были получены двумя путями: на основании сравнения суммарных каталитических активностей, на основании сравнения типа и состава образующихся продуктов. [5]
Важным свидетельством в пользу того, что реакция ( IV) представляет собою ключевую реакцию в процессе каталитического разложения перекиси, служит то, что скорость образования первичного комплекса с перекисью водорода является достаточно большой, чтобы объяснить суммарную каталитическую активность, если механизм реакции такой, что процесс может протекать быстро. [6]
Тогда, если в тетраплоидной клетке находятся в одинаковых соотношениях две произвольных аллеля из четырех, доля гибридных белков в клетке составит [ 1 - 2 ( 1 / 2) л ] от всех имеющихся, где п - муль-тимерия белка. При одновременном нахождении в клетке четырех аллелей доля гибридных ферментов составит [ 1 - 4 ( 1 / 4) ], т.е. будет большей, чем в первом случае, а значит и суммарная активность этого фермента тоже будет более значительной. Таким образом, изменяя соотношение и количество аллелей, можно менять суммарную каталитическую активность клетки. С таким явлением, может быть, исследователи столкнулись при изучении комплементации у полиплоидных дрожжей. В работе [9], где изучалась комплементация у трипло-идов, восстановление нормы у триплоидов при объединении в клетке трех аллелей, попарно некомплементарных в диплоиде, можно объяснить, в частности, варьированием соотношения аллелей или, что то же самое, изменением концентраций полипептицных цепей. [7]
Страницы: 1