Гидроксилаз
Cтраница 3
Большинство микроорганизмов имеет тот или иной специфический набор гидроксилирующих ферментов - стероид-гидроксилаз, каждый из которых осуществляет гидроксилирование, как правило, лишь в одно положение. При этом следует, во-первых, подчеркнуть, что отдельные гидроксилазы также индивидуальны у каждого микроорганизма и различаются в ряде аспектов; поэтому то, что справедливо для одной гидроксилазы, может быть неприменимо к другой. Во-вторых, один и тот же фермент может, хотя и очень редко, в зависимости от строения субстрата, гидроксилировать его в различные положения. Примером может служить Fusarium lini, гидроксилаза которого вводит в C / D-пгранс-стероиды 15а - оксигруппу, а в C / D-tyuc - стероиды - 12 ( 3-оксигруппу. [31]
В нормальных физиологических условиях большая часть фенилаланина превращается в тирозин. Фермент, ответственный за эту реакцию - фенил-аланин-гидроксилаза - напоминает другие типичные гидроксилазы ( см. гл. XV) тем, что в реакцию вовлекаются молекулярный кислород и восстановленный НАДФ. Этот фермент локализуется в микросомной фракции клетки. [32]
Аналогичное предположение было выдвинуто и доказано при сравнении гидроксилирования C / D-цис - и С / В-трамс-стероидов. Возникает вопрос, образует ли микроорганизм два специфических гидроксилирую-щих фермента - 12р - и 15а - гидроксилазы, или же дело идет только об одном относительно неспецифичном ферменте. [33]
Однако их опыты показали, что при этом пороке обмена из меченого фенилаланина все же образуется некоторое количество тирозина. Указанные авторы полагают, что результаты их исследования могут быть объяснены а) уменьшением ( но не полным отсутствием) гидроксилазы фенилаланина ( стр. [34]
Большинство микроорганизмов имеет тот или иной специфический набор гидроксилирующих ферментов - стероид-гидроксилаз, каждый из которых осуществляет гидроксилирование, как правило, лишь в одно положение. При этом следует, во-первых, подчеркнуть, что отдельные гидроксилазы также индивидуальны у каждого микроорганизма и различаются в ряде аспектов; поэтому то, что справедливо для одной гидроксилазы, может быть неприменимо к другой. Во-вторых, один и тот же фермент может, хотя и очень редко, в зависимости от строения субстрата, гидроксилировать его в различные положения. Примером может служить Fusarium lini, гидроксилаза которого вводит в C / D-пгранс-стероиды 15а - оксигруппу, а в C / D-tyuc - стероиды - 12 ( 3-оксигруппу. [35]
 |
Энергетический профиль реакции гидратации фумаровой кислоты до яблочной кислоты под действием фумаразы. [36] |
В зависимости от того, какие реакции катализируют ферменты, их подразделяют на шесть основных групп. Оксидоредуктазы, катализируют окислительно-восстановительные процессы. К ним принадлежат дегидрогеназы, оксидазы, ре-дуктазы, трансгидрогеназы и гидроксилазы. [37]
Фенилаланин относится к незаменимым аминокислотам, поскольку ткани животных не обладают способностью синтезировать его бензольное кольцо. В то же время тирозин полностью заменим при достаточном поступлении фенилаланина с пищей. Реакция гидроксилирования катализируется специфической фенилаланин-4 - монооксигеназой, которая в качестве кофермента содержит, как все другие гидроксилазы, тетрагидро-биоптерин. В процессе трансаминирования тирозин превращается в и-оксифенилпировиноградную кислоту, которая под действием специфической оксидазы подвергается окислению, декарбоксилированию, гидро-ксилированию и внутримолекулярному перемещению боковой цепи с образованием гомогентизиновой кислоты; эта реакция требует присутствия аскорбиновой кислоты, роль которой пока не выяснена. Дальнейшее превращение гомогентизиновой кислоты в малеилацетоуксусную кислоту катализируется оксидазой гомогентизиновой кислоты. Малеилацетоуксус-ная кислота под действием специфической изомеразы в присутствии глу-татиона превращается в фумарилацетоуксусную кислоту, подвергающуюся гидролизу с образованием фумаровой и ацетоуксусной кислот, дальнейшие превращения которых уже известны. [38]
 |
Пути метаболизма катехоламинов ( основной путь выделен. [39] |
Нук-леотиды и катехоламины транспортируются в везикулы отдельными носителями под действием градиента рН, генерируемого электрогенным АТРазным насосом. В качестве высокомолекулярных компонентов хромаффинных гранул идентифицирована группа кислых секреторных белков хромогранинов и меньшие количества допамин - - гидроксилазы - главного компонента везикулярной мембраны. Как растворимый секреторный, так и мембранно-связанный белок содержат мукополисахариды. [40]
Микробиологическое гидроксилирование ароматического кольца широко освещено в литературе, однако в настоящее время этот метод имеет - гораздо меньшую синтетическую ценность для химика-органика, чем гидроксилирование алифатических соединений, которому посвящено значительно меньшее число работ. Это обусловлено по крайней мере двумя причинами: во-первых, гидроксилированное ароматическое кольцо становится чрезвычайно чувствительным к дальнейшему окислению с помощью других ферментов в клетке или в бесклеточных экстрактах, которое протекает обычно с разрывом кольца и разрушением его до меньших фрагментов ( это будет обсуждаться в гл. Однако известные, пусть малочисленные более или менее полезные синтетические методы позволяют надеяться на то, что в этой области будет еще многое открыто. Выделение первой гидроксилазы в кристаллическом виде [13] позволяет предполагать, что процесс последовательной деградации можно будет в конечном счете расчленить на индивидуальные реакции. [41]
Оксигена-ция протекает как атака субстрата образующимся супероксид-анионом кислорода. Одной из биологически важных реакций такого типа является превращение 3-каро-тина в витамин А. Монооксигеназы требуют участия в реакции НАДФН, атомы водорода которого взаимодействуют с одним из атомов кислорода, поскольку только один электрон связывается с субстратом. К широко распространенным монооксигеназам относятся разнообразные гидроксилазы. Они принимают участие в окислении аминокислот, оксикислот, полиизопреноидов. [42]
В животных тканях быстрый синтез ферментов наблюдается реже. Механизм его ( индуцирующий синтез) изучен только для небольшого числа ферментов: тирозинтрансаминазы, серии - и треониндегидратазы, триптофанпирролазы и др. - в ответ на введение гормонов или прием белковой пищи. Однако при поступлении в организм некоторых ядов, канцерогенных веществ, алкалоидов, инсектицидов через несколько дней наблюдается резкое повышение активности ( соответственно количества) ферментов-гидроксилаз ( монооксигеназ) эндоплазматической сети клеток печени, окисляющих чужеродные вещества в нетоксичные для организма продукты. Описаны случаи, когда под действием подобных гидроксилаз чужеродные вещества превращаются в организме в более токсичные соединения. [43]
Значение витамина D начинает проясняться в последнее время. Получены доказательства, что при физиологических условиях кальциферолы функционально инертны. D выполняет свои биологические функции в организме в форме образующихся из него активных метаболитов, в частности 1 25-диоксихолекальциферола [ сокращенно обозначается 1 25 ( OH) 2D3 ] и 24 25-диоксихолекальциферола [ 24 25 ( OH) 2D3 ], причем если гидрокси-лирование в 25 - м положении осуществляется в печени, то этот процесс в 1 - м положении протекает в почках. Ферменты, катализирующие эти реакции, называются гидроксилазами, или монооксигеназами. В реакциях гидрокси-лирования используется молекулярный кислород. Показано, что специфическая la - гидроксилаза содержится, помимо почек, в костной ткани и плаценте. Имеются бесспорные доказательства, что именно эти активные метаболиты, выполняя скорее гормональную, чем биокаталитическую, роль, функционируют в системе гомеостатической регуляции обмена кальция и минерализации костной ткани. В частности, 1 25 ( OH) 2D3 участвует в регуляции процессов всасывания Са и Р в кишечнике, резорбции костной ткани и реабсорбции Са и Р в почечных канальцах. Методом ауторадиографии показано накопление 1 25 ( OH) 2D3 в ядрах клеток органов-мишеней ( почки, мозг, поджелудочная железа, гипофиз, молочная железа), где он способствует синтезу мРНК, Са-связывающих белков и гормонов, регулирующих обмен кальция; в то же время он не обнаруживается в печени, селезенке, скелетной и сердечной мышцах. [44]
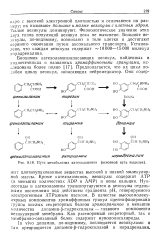
Образующиеся биогенные амины - триптамин, серотонин, дофамин обладают сильным фармакологическим действием на множество физиологических функций человека и животных. Так, триптамин и серотонин оказывают сосудосуживающее действие. Кроме этого, серотонин участвует в регуляции артериального давления, температуры тела, дыхания и почечной фильтрации, является нейромедиатором, который вызывает изменение поведения, например при шизофрении. Дофамин, возможно, сам является нейромедиатором, а также предшественником широко известного медиатора норэпинефрина и гормона адреналина. Источником ДОФА в организме является тирозин, который под действием специфической гидроксилазы превращается в 3 4-диоксифенилаланин. Тирозингидроксилаза открыта в надпочечниках, в тканях мозга и периферической нервной системы. [45]
Страницы: 1 2 3 4