Гидролиз - молекула
Cтраница 3
Для реакции первого порядка эта зависимость должна выражаться прямой линией. Фактическая же форма кривой указывает на возможность некоторого гидролиза молекул аппрета наряду с повышением его сопротивления десорбции по мере экстрагирования. Это не противоречит экспериментальным данным Шрейдера и др. [9], согласно которым адсорбированный слой аппрета обладает неодинаковой стойкостью к гидролитической десорбции. [31]
 |
Схема строения тонкой нити мышечного волокна. [32] |
Мышцы способны использовать для сокращения около 30 % химической энергии, запасенной в молекулах АТФ. Они показали, что источником энергии сокращения мышц является гидролиз молекул АТФ при взаимодействии с актомиозином. [33]
В третьей группе катионов, по-видимому, лимитирующей стадией является гидролиз молекулы воды в координационной оболочке. Скорости в этой группе сравнительно низкие и меняются для различных лигандов. Примерами таких катионов служат Fe3 ( стр. Константы скорости для реакций Fe3 с различными анионами изменяются в значительной степени параллельно с основностью аниона ( стр. [34]
Большие отрицательные величины стандартных свободных анергий гидролиза обусловлены энергией гидратации отрицательно заряженных продуктов гидролиза и перестройкой их электронных оболочек. Из табл. 8 следует, что значение стандартной свободной энергии гидролиза молекулы АТФ занимает промежуточное положение между высокоэнергетическими ( фосфоэнолпиру-ат) и низкоэнергетическими ( глюкозо-6 - фосфат) соединениями. Это одна из причин того, что молекула АТФ является удобным универсальным переносчиком фосфатных групп. [35]
 |
Процесс терминации синтеза белка ( схема. [36] |
АТФ до 2 молекул АДФ, поскольку пирофосфат подвергается распаду на 2 молекулы неорганического фосфата. Для включения амино-ацил - тРНК в аминоацильный центр используется энергия гидролиза молекулы ГТФ на ГДФ и неорганический фосфат. Наконец, транслокация транслирующей 70S рибосомы также нуждается в энергии гидролиза еще одной молекулы ГТФ. Таким образом, энергетические потребности синтеза каждой пептидной связи эквивалентны энергии гидролиза 2 молекул АТФ и 2 молекул ГТФ ( т.е. гидролиз четырех макроэргических фосфатных связей) до соответствующих нуклеозиддифосфатов. Легко представить, насколько велики энерготраты каждой клетки при синтезе не только одной молекулы белка, а множества молекул самых разнообразных белков в единицу времени. [37]
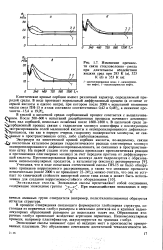 |
Изменение прочности связи стекловолокно-смола при длительном воздействии жидких сред при 293 К ( а, 323 К ( 6 и 353 К ( в. [38] |
В кислой и щелочной средах сорбционный процесс сочетается с выщелачива - О нием. В щелочной среде де - Ч сорбционный процесс связан с гидролизом молекул олигоэфира с последующим экстрагированием солей малеиновой кислоты и оксипропилированного дифенилол - Л пропанола, так как в первую очередь гидролизу подвергаются молекулы, не свя-ные в пространственную сетку, либо слабоупакованные. [39]
С другой стороны, в мягких, легко деформируемых цепочках эти возбуждения существенно отличаются друг от друга. Имеются все основания считать, что в а-спиральных белковых молекулах при гидролизе молекул АТФ должны образовываться солито-ны. Именно солитоны могут весьма эффективно переносить энергию возбуждения на сравнительно большие расстояния. [40]
Такое состояние мембраны называется энергизированным. Оно обеспечивается за счет энергии переноса электронов при окислении некоторых веществ или гидролиза молекул АТФ. Хотя молекулярный механизм этих процессов совершенно не установлен, использование гипотезы Митчелла оказывается полезным для интерпретации ряда явлений. [41]
I, III, VIII и XIV содержание аспарагиновой и глютамино-вой кислот, глицина, аланина, лейцина, тирозина, гистамина, лизина, аргинина и триптофана было ( в пределах ошибки опыта одним и тем же. Некоторые отклонения в количествах других аминокислот, вероятно, связаны с различием глубины гидролиза молекулы антитела, скоростью отщепления и разделения аминокислот. [42]
Разрыв связей между тонкими нитями и головами миозиновы. АТФ и ионами Mg2 n мышцах, не приводит, однако, к гидролизу молекул АТФ. Комплексы Mg АТФ - присоединяются к головам миозиновых молекул. Такое существенно различное поведение топких нитей в саркомере и F-актиновых нитей без молекул тропонина и тропомиозина в растворе с миозиновыми молекулами обусловлено наличием в тонких нитях кроме молекул актина двух других белков: тропомиоаина и тропонина. [43]
Внутри волокна потенциал отрицателен по отношению к внешней среде, потенциал которой обычно принимается равным нулю. Разность концентраций ионов и потенциалов обусловлена работой ионных насосов, выбрасывающих за счет энергии гидролиза молекул АТФ три иона Na из внутренней области аксона и впускающих два иона К внутрь волокна. [44]
Процесс разрушения большой молекулы крахмала под действием амилазы протекает значительно сложнее, чем это предполагалось ранее. Так как а-амилаза способна разрывать цепочку полисахарида в любом месте, то при известной степени гидролиза молекулы крахмала может образоваться не только мальтоза, но и глюкоза. Это предположение подтверждается опытами по осахариванию крахмала препаратом Asp. [45]
Страницы: 1 2 3 4