Хемиосмотическая гипотеза
Cтраница 2
Сколько ионов Н содержится в одной митохондрии. Хемиосмотическая гипотеза предполагает, что в результате переноса электронов ионы Н выталкиваются из матрикса митохондрий наружу, вследствие чего между двумя сторонами мито-хондриальной мембраны возникает градиент рН, при котором наружная фаза оказывается более кислой, чем внутренняя. Способность ионов Н диффундировать в обратном направлении, из окружающей среды в митохондриальный ма-трикс ( где их концентрация ниже), служит, согласно этой гипотезе, движущей силой для синтеза АТР, катализируемого FoF AT / Разой. В митохондриях, суспендированных в среде с рН 7 4, происходит окислительное фосфорилирование. [16]
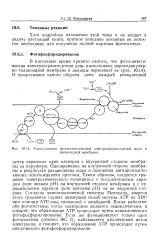 |
Расположение фотосиитетической электроитранспортной цепи в. [17] |
Одновременно на внутренней стороне мембраны в результате расщепления воды и окисления дигидропласто-хинона освобождаются протоны, Таким образом, создается градиент электрохимического потенциала с положительным зарядом ( или низким рН) на внутренней стороне мембраны и с отрицательным зарядом ( или высоким рН) на ее наружной стороне. Согласно хемиосмотической гипотезе Митчелла, энергия этого градиента служит движущей силой синтеза АТР из ADP при помощи АТР-азы, связанной с мембраной. Когда работают обе фотосистемы, поток электронов является нециклическим, и говорят, что образование АТР происходит путем нециклического фотофосфорилирования. Если же функционирует только одна фотосистема ( обычно ФС I), наблюдается циклический поток электронов, и говорят, что образование АТР происходит путем циклического фотофосфорилирования. [18]
Окислительное ( расформирование в инвертированных субмитохондриалъных пузырьках. Согласно хемиосмотической гипотезе, во время переноса электронов из интактных митохондрий откачиваются наружу ионы Н, что приводит к возникновению градиента рН между двумя сторонами митохондриальной мембраны. Этот градиент рН заключает в себе энергию, благодаря которой ионы Н перемещаются в обратном направлении-из окружающей среды в митохондриальный матрикс. Удалось показать, что полученные из внутренней митохондриальной мембраны инвертированные пузырьки, у которых FoFj-АТРазные головки обращены наружу ( рис. 17 - 15), тоже способны к окислительному фосфорилированию. [19]

Под влиянием индуцированного светом потока электронов возникает, следовательно, трансмембранный градиент концентрации ионов Н, так что с внутренней стороны тилакоидных пузырьков среда становится более кислой, чем с наружной. Все эти свойства согласуются с хемиосмотической гипотезой, предложенной первоначально для объяснения ркислительного фосфорилирова-ния, а позднее распространенной и на фотосинтетическое фосфорилирование. Схема на рис. 23 - 16 изображает поток ионов Н, движимых световой энергией; ионы Н 1 поступают из стромы внутрь тилакоида и вновь возвращаются наружу через молекулы АТР-синтетазы. В 1966 г. Андрэ Ягендорф поставил важный эксперимент, доказавший, что источником энергии для синтеза АТР действительно может служить трансмембранный градиент рН, обращенный щелочным концом наружу. [21]
Итак, хемиосмотическая гипотеза не требует никакого высокоэнергетического химического агента, который в качестве общего промежуточного продукта обеспечивал бы сопряжение между переносом электронов и синтезом АТР. Переносчиком энергии, связывающим два эти процесса, служит, согласно хемиосмотической гипотезе, градиент концентрации ионов Н между двумя сторонами митохондриальной мембраны. [22]

Когда этот градиент начинал уменьшаться вследствие выхода ионов Н из тилакоидов в среду, происходило образование АТР из ADP и фосфата. Поскольку АТР синтезировался в темноте, этот эксперимент доказывал, что трансмембранный градиент рН есть высокоэнергетическое состояние, посредством которого энергия переноса электронов передается АТР-синтетазе, получающей таким образом возможность генерировать АТР, как это постулируется хемиосмотической гипотезой. [24]
Согласно этой гипотезе, функция переноса электронов, происходящего во внутренней митохондриальной мембране, заключается в том, чтобы откачивать ионы Н из матрикса митохондрии в наружную среду и таким путем создавать между двумя водными фазами, которые разделяет эта мембрана, градиент концентрации ионов Н с более кислым значением рН снаружи. Такой градиент, при котором концентрация ионов Н снаружи выше, чем внутри митохондрии, обладает потенциальной энергией ( разд. Хемиосмотическая гипотеза постулирует далее, что ионы Н, выведенные наружу за счет энергии переноса электронов, снова устремляются внутрь, в митохондриальный матрикс, через специальные каналы, или поры, для этих ионов в молекулах FgFj-АТРазы. В этом случае они перемещаются по градиенту концентрации и во время их перехода через молекулы АТРазы выделяется свободная энергия. Именно эта энергия и служит движущей силой для сопряженного синтеза АТР из ADP и фосфата. [25]
Хемиосмотическая гипотеза, сформулированная английским биохимиком Питером Митчеллом, исходит из совершенно иного, нового принципа. Постулируется, что перенос электронов сопровождается выкачиванием ионов Н из матрикса через внутреннюю митохон-дриальную мембрану в наружную водную среду. Синтез АТР, требующий затраты энергии, осуществляется именно за счет осмотической энергии, присущей этому градиенту. Можно думать, что именно хемиосмоти-ческая теория наиболее точно отражает организующий принцип окислительного фосфорилирования. Рассмотрим некоторые характерные особенности этого процесса, свидетельствующие в пользу хемиосмотической гипотезы. [26]
В работах лабораторий Либермана и Скулачева расположение дыхательной цепи определялось по ее способности образовывать мембранный потенциал. В среду вводились различные доноры и Акцепторы электронов, не проникающие сквозь мембрану. Оказалось, что эти вещества взаимодействуют лишь с цитохромом с в митохондриях. Установлено, что транспорт протонов и ( или) электронов по дыхательной цепи действительно происходит. В других экспериментах определена локализация компонентов в мембране митохондрий. На рис. 13.10 показано вероятное расположение цепи. Согласно хемиосмотической гипотезе, любая со-тфягающая система должна создавать электрохимический потенциал ионов Hf. Действительно, опыты с проникающими синтетическими ионами показали возникновение Дг з в митохондриях, СМЧ, хлоропластах ( см. гл. В то же время теория Митчелла встречается с трудностями и вызывает возражения. Блюменфельд приводит аргументы, показывающие невозможность построения машины Митчелла в конденсированной фазе. В такой машине АТФ-синтетаза использует разность концентраций протонов в водной фазе по обе стороны мембраны для выполнения внешней работы. [27]
Страницы: 1 2