Гистон
Cтраница 4
Протаминыи гистоны являются наиболее простыми белками. Они отличаются от других белков тем, что имеют слабощелочной характер. Вследствие этого, первые исследователи рассматривали протамины и гистоны не как белковые вещества, а как особые органические основания, близкие к растительным алкалоидам. Впоследствии стало известно, что щелочной характер этих белков обусловлен тем, что в состав их молекулы входят преимущественно аминокислоты - л изин, аргинин и г и с т и д и н, обладающие. Количество содержащихся в протаминах диаминокислот доходит до 80 % и более. [46]
Поскольку гистоны являются важной составной частью хроматина, следует уделить некоторое внимание их происхождению и биологии. Хотя в интерфазном ядре синтез гистонов обычно идет параллельно репликации ДНК, эти два процесса не обязательно должны быть связаны друг с другом. Синтезированные гистоны каким-то образом ( пока неизвестно каким) перемещаются из ядрышка в хроматин и соединяются там с соответствующими частями геномной ДНК. Новый гистон, поступающий из ядрышка, в котором он был синтезирован, должен покрывать образующийся дефицит гистонов. Механизмы контроля и регулирования такого транспорта гистонов и их распределения требуют дальнейшего изучения. Гистоны, распределенные на хроматине, в отличие от ДНК не являются метаболически инертными, но обменные процессы, в которых они участвуют, протекают медленно. [47]
Кроме гистонов в хроматине присутствует большое количество различных негистоновых белков, характер взаимодействия которых с нуклеосомной ДНК пока не ясен. Наиболее богато представлены негистоновые белки HMG 14 и 17, функция которых остается все еще не изученной. HMG 14 и 17 - это близкие по структуре белки, несущие большое количество заряженных групп. Они состоят соответственно из 68 и 74 аминокислотных остатков. HMG 14 и 17 связываются с обращенной внутрь нуклеосомы стороной двойной спирали ДНК и не меняют существенным образом общую форму нуклеосомы. Создается впечатление, что эти два белка занимают свободную внутреннюю область ДНК нуклеосомы. [48]
АДФ-рибозилирование гистонов меняет их структуру и способность взаимодействовать с хроматином [ Poirier G. G., Savad P. Помимо гистонов, поли - АДФ-рибозилированию подвергаются и неги-стоновые белки хроматина, что особенно характерно для быстрорастущих тканей. В качестве акцепторов АДФ-рибозы могут выступать также белки HMG-1 и HMG-14 хроматина. АДФ-рибозилирование гистона HI приводит к образованию димеров, состоящих из двух молекул гистона, которые связаны цепью поли - АДФ-рибозы. Эта цепь включает 15 - 16 мономерных единиц. Хроматин в таком комплексе находится в сильно конденсированном состоянии, что обусловливает его устойчивость к микрококковой нук-леазе. [49]
Связь гистонов с оксиапатитом имеет, очевидно, катионный характер, что позволяет фракционировать их ( и одновременно диссоциировать от ДНК) при ступенчато увеличивающейся концентрации NaCl. В 0 1 М фосфатном буфере ДНК остается связанной с. [50]
Кроме гистонов в хроматине присутствует большое количество различных негистоновых белков, характер взаимодействия которых с нуклеосомной ДНК пока не ясен. Наиболее богато представлены негистоновые белки HMG 14 и 17, функция которых остается все еще не изученной. HMG 14 и 17 - это близкие по структуре белки, несущие большое количество заряженных групп. Они состоят соответственно из 68 и 74 аминокислотных остатков. HMG 14 и 17 связываются с обращенной внутрь нуклеосомы стороной двойной спирали ДНК и не меняют существенным образом общую форму нуклеосомы. Создается впечатление, что эти два белка занимают свободную внутреннюю область ДНК нуклеосомы. [51]
Кроме гистонов в хроматине присутствует большое количество различных негистоновых белков, характер взаимодействия которых с нуклеосомной ДНК пока не ясен. Наиболее богато представлены негистоновые белки H. HMG 14 и 17 - это близкие по структуре белки, несущие большое количество заряженных групп. Они состоят соответственно из 68 и 74 аминокислотных остатков. HMG 14 и 17 связываются с обращенной внутрь нуклеосомы стороной двойной спирали ДНК и не меняют существенным образом общую форму нуклеосомы. Создается впечатление, что эти два белка занимают свободную внутреннюю область ДНК нуклеосомы. [52]
Строение гистонов несколько сложнее: в их состав входит тирозин, но триптофан содержится в небольших количествах или отсутствует. [53]
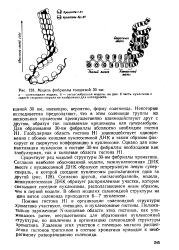 |
Модель фибриллы толщиной 30 нм. [54] |
Помимо гистона HI в организации соленоидной структуры хроматина участвуют, очевидно, и нуклеосомные гистоны. Положительно заряженные N-концевые области этих гистонов, как упоминалось ранее, несущественны для образования нуклеосомной структуры, но вовлечены в организацию соленоидной структуры хроматина. Удаление этих участков с помощью мягкого расщепления гнстонов трипсином в составе хроматина приводит к необратимому разворачиванию соленоида. [55]
Страницы: 1 2 3 4