Гидродинамические данные
Cтраница 2
Анализ данных радиационных наблюдений показал, что для участка ЛСК Радон характерны неоднозначность гидродинамической ситуации, наличие локальных потоков, связанных, видимо, с дренажными траншеями и канавами. Поэтому, используя региональные гидродинамические данные, достаточно сложно описать процесс распространения загрязнения на участке ЛСК. На некотором удалении от зданий 668, 668А, 668Б, 57 в большинстве скважин регистрируются пики активности радионуклидов, которые могут быть связаны с разновременными утечками из различных источников. К 1994 - 1996 гг. в этих скважинах также наблюдается относительная стабилизация удельных активностей радионуклидов, которые, тем не менее, остаются в основном на 2 - 3 порядка выше фоновых значений. Кроме того, по режимным радиационным наблюдениям выделяются скважины, где отсутствуют ярко выраженные пики загрязнения, удельные активности стронция и трития меняются по данным скважинам хаотично. Расположены они, как правило, в направлениях, противоположных или перпендикулярных региональному потоку от мест предполагаемых утечек. Однако скачкообразные превышения фоновой р-активности ( вплоть до последних лет иногда на порядок - скважины 1А, 6) заставляют предположить, что и в этих скважинах ( З - активность повышена именно за счет радиостронция. [16]
Это уравнение для Рг 30 и в пределах Re от 10000 до 50000 ( в которых справедливо уравнение Блазиуса) дает достаточно хорошее совпадение с экспериментальными результатами. Так как это уравнение основывается на гидродинамических данных, а не на тепловых, то оно подтверждает аналогию между теплоотдачей и трением. [17]
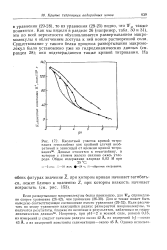
Как мы видели в разделе 26 ( например, табл. 30 и 31), это по всей вероятности обусловливается развертыванием макромолекулы и облегчением доступа к ней ионов растворенной соли. Существование у такого белка процесса развертывания макромолекул было установлено уже из гидродинамических данных ( см. раздел 28); они подтверждаются также кривой титрования. [19]
Вследствие гидратации полярных групп мицеллообра-зующих молекул ( ионов) на поверхности мицелл имеется гидратная оболочка. Количество связанной воды на поверхности мицелл может быть определено на основании гидродинамических данных: по вязкости, скорости диффузии, скорости седиментации в ультрацентрифуге. [20]
Электронная микрофотография указывает ( см. рис. 37), что молекула состоит из трех более или менее сферических шариков; диаметром 65, 50 и 65 А соответственно соединенных тонкими нитями диаметром 15 А или меньше. Общая длина составляет 475 А. Гидродинамические данные, с другой стороны, указывают на продолговатый эллипсоид с длиной около 700 А и максимальным диаметром 38 А. [21]
С точки зрения математического описания экстракция является сложнейшим многофакторным процессом. Факторы, определяющие механизм протекания и эффективность этого технологического процесса, с достаточной полнотой отражены в предыдущих главах. Сложность математических моделей, отображающих характеристики промышленного процесса экстракции, обусловлена тем, что в них фокусируется вся совокупность равновесных, кинетических и гидродинамических данных, получаемых на разных стадиях изучения процесса: от лабораторных исследований до промышленных испытаний. [22]
Молекулярная масса углевод-белковых полимеров является величиной усредненной, учитывающей полидисперсность полимеров. Уже на стадии выделения, очистки и определения гомогенности этих биополимеров можно приблизительно оценить их свойства и молекулярную массу. Предварительное определение вязкости веществ и содержания в них сиаловых кислот позволяет выбрать те или иные пути дальнейшего исследования. Молекулярная масса углевод-белковых полимеров может быть рассчитана на основании гидродинамических данных, коэффициентов седиментации, диффузии, характеристической вязкости. [23]
Поскольку гидратационная вода должна увеличивать диаметр стержня в большей степени, чем его длину, следует ожидать, что отношение осей для гидродинамического стержня будет меньшим, чем для реальной частицы. Однако зависимость функций v и F от отношения осей такова, что уменьшение величины а / b в результате гидратации почти полностью компенсируется увеличением гидродинамического объема. Благодаря такой компенсации значения а, р и у для длинных палочкообразных молекул, полученные с помощью гидродинамических данных, близки к значениям, рассчитанным на основании рентгенограмм и электронных микрофотографий. [24]
При сравнении этих данных мы должны учитывать возможность того, что электронная микрофотография является обманчивой. Может быть, что замороженная и высушенная молекула, которую видно в электронный микроскоп, отличается от молекулы в растворе. Возможно также и то, что нити, которые соединяют шарики ( и которые плохо видны), простираются за пределы крайних шариков, так что истинная молекулярная длина превышает размер 475 А, измеренный между внешними кромками шариков. Сомнения относительно точности данных, получаемых при помощи электронного микроскопа, яляются, однако, неуместными в отношении главного обстоятельства, которое выявляется при сравнении двух результатов. Это обстоятельство состоит в том, что размеры, приведенные на рис. 37, являются абсолютно возможными и не несовместимыми с гидродинамическими данными. Поскольку уравнения для интерпретации гидродинамических данных являются ограниченными в том смысле, что они требуют использования модели вытянутого эллипсоида для всех жестких частиц, из этого следует, что мы можем вычислять по гидродинамическим данным только размеры эллипсоида, наиболее точно соответствующего действительной молекуле. Различие между лучшим эллипсоидом для фибриногена и формой, данной на рис. 37, должно рассматриваться как мера неточности, которая относится ко всем формам, выведенным из гидродинамических измерений, даже когда они комбинируются с данными по рассеянию света. [25]
Страницы: 1 2