Палеонтологические данные
Cтраница 1
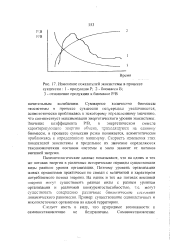 |
Изменение показателей экосистемы в процессе сукцессии. I - продукция Р. 2 - биомасса В. 3 - отношение продукции к биомассе Р / В. [1] |
Палеонтологические данные показывают, что на одних и тех же потоках энергии в различные исторические периоды существовали виды разного уровня организации. Поэтому уровень организации живых организмов практически не связан с величиной и характером потребляемого потока энергии. На одних и тех же потоках внешней энергии могут существовать разные виды с разным уровнем организации и различной конкурентоспособностью, т.е. могут существовать совершенно различные биологические состояния динамического равновесия. [2]
Палеонтологические данные показывают, что предки современного человека занимали нишу собирателя - вида с широким спектром потребления растительной и примерно в 10 раз меньше доступной животной пищи. [3]
Палеонтологические данные показывают, что предки современного человека, жившие в эпоху раннего палеолита, занимали экологическую нишу собирателя - вида с широким потреблением растительной и примерно в 10 раз менее доступной животной пищи. [4]
Геологические и палеонтологические данные решительно противоречат этому. [5]
Анализ геологических и палеонтологических данных показывает, что наша планета облучалась столь же интенсивной солнечной радиацией, как и сейчас, по крайней мере 500 миллионов лет. На основе астрономических данных можно сделать заключение, что, вероятно, с Солнцем практически не происходило никаких заметных изменений в течение около 5 миллиардов лет, то есть со времени образования Солнечной системы. Эти факты, а также примерная масса Солнца, известная ученым еще в XIX веке, вызвали различные гипотезы о природе солнечной энергии. Достаточно произвести несложные подсчеты, чтобы убедиться, что источником этой энергии не могут быть химические реакции, происходящие при сгорании таких обычных горючих, как уголь или нефть. [6]
Одних лишь палеонтологических данных недостаточно для обоснования того, что эволюция действительно происходила, однако они подтверждают факт прогрессивного возрастания сложности организмов. Ископаемые остатки были хорошо известны еще до того, как эволюция получила всеобщее признание. [7]
С учетом кернового материала и палеонтологических данных, привязанных к материалам каротажа, битуминозные отложения были расчленены на отдельные части, которые впоследствии были переведены в ранг подсвит и пачек. Территории, на которых отмечается одинаковое строение разреза объединялись в районы, которым были присвоены географические наименования. Баженовская свита развита в Тамбейско-Омской зоне. Верхний контакт с перекрывающими образованиями повсеместно согласный, нижняя граница резкая. В сводах некоторых поднятий георгиевская свита выпадает из разреза и битуминозные осадки несогласно залегают на породах верхневасюганской подсвиты. Малая мощность битуминозных пород связана с активным ростом Нижневартовского свода в этот отрезок времени. [8]
Строение неокома рассмотрено исходя из клиноформной модели, предложенной А.Л. Наумовым, приведены палеонтологические данные ( находки остатков архистратиграфических групп организмов), в целом подтверждающие клиноформное строение неокома. Рассмотрены ограничения биостратиграфического метода при изучении разнофациальных, накапливавшихся с лавинными скоростями неокомских отложений. [9]
Холм Юдика и соседний холм Скальпелло представляются как серия разобщенных чешуи, несмотря на противоположное утверждение, на основании геофизических и палеонтологических данных. [10]
Среднеюрские отложения, широко развитые в Прикумской нефтегазоносной зоне, трансгрессивно перекрывают нижнеюрские, а в зонах размыва последних ложатся на карбоновые сланцы или на пермо-триас. Литолого-фациальные и палеонтологические данные позволяют выделить байосский и условно батский ярусы, в которых выделяются два песчаных пласта. Нижняя песчаная пачка, именуемая V пластом, так называемая величаевская свита, по возрасту относится к нижнему байосу. V пласт - наиболее проницаемая часть песчаных разрезов, четко выделяемая по электрокаротажной характеристике по отношению к вмещающим глинам. [11]
В результате этих работ была предложена новая, более детальная стратиграфическая схема, основанная на сочетании палеонтологических данных с принципом цикличности в осадконакоплении. [12]
Получается, что ЭМП являются необходимым условием зарождения жизни на Земле. И их роль не ограничивается ролью акушера, помогающего только при родах. Недавние палеонтологические данные свидетельствуют о том, что периоды инверсии геомагнитных полюсов сопровождаются изменением видового состава донной морской фауны. [13]
Таким образом, первым этапом палеогеографической реконструкции является восстановление условий осадконакопления. Состав осадков и палеонтологические данные могут быть использованы для определения климата. Например, наличие карбонатных пород с широким развитием колониальных кораллов в качестве породообразовате-лей свидетельствует об определенной морской обстановке и теплом климате, наличие бокситов латеритного типа и угленосных осадков с теплолюбивой и влаголюбивой флорой - об определенных континентальных обстановках и теплом и влажном климате. [14]
Все эти науки оказывают большую помощь при определении таксономических связей. Наука о распространении животных - зоогеография, или хорология, заняла, наконец, подобающее ей место в систематике. Безусловно, палеонтологические данные очень облегчают любое таксономическое исследование, и Аркелл и Мой-Томас [40] составили соответствующую сводку Палеонтология и таксономическая проблема. Влияние климата и погоды на насекомых имеет также большое значение и может существенно помочь в том или ином конкретном таксономическом исследовании. Выдающейся является работа Уварова Насекомые и климат [2155], и с ней должен детально ознакомиться каждый изучающий эту науку. [15]
Страницы: 1 2