Движение - радикал
Cтраница 2
Итроксилы полезными в качестве меток и зондов, это анизотропия их магнитных параметров, благодаря которой радикалы, различным образом ориентированные в образце, обладают разными резонансными частотами ЭПР. Реориентационные движения радикала приводят к частотному обмену и вызывают в спектре релаксационные изменения, анализ которых дает информацию о характере и частотах движений. Наконец, воздействие окружения на нитр-оксил проявляется в изменении величин собственно магнитных параметров ( компонент g - и Л - тензоров), которые, следовательно, могут быть использованы для характеристики этого воздействия. [16]
Продольный Процесс Т является истинным процессом релаксации энергии. Oil происходит вследствие того, что движение радикала вызывает флуктуации локальных магнитных полей, а эти флуктуации могут стимулировать электроны к переходу в более низкое энергетическое состояние с переносом избытка энергии к решетке. Эффективность процесса определяется флуктуациями, и поэтому изучение ширины линий является источником ценной информации о движении молекул в жидкостях. Изучение ширины линий может показать, как быстро вращаются молекулы и происходит ли вращение преимущественно покрут одной оси радикала. [17]
Продольный процесс Г, является истинным процессом релаксации энергии. Он происходит вследствие того, что движение радикала вызывает флуктуации локальных магнитных полей, а эти флуктуации могут стимулировать электроны к переходу в более низкое энергетическое состояние с переносом избытка энергии к решетке. Эффективность процесса определяется флуктуациями, и поэтому изучение ширины линий является источником ценной информации о движении молекул в жидкостях. Изучение ширины линий может показать, как быстро вращаются молекулы и происходит ли вращение преимущественно вокруг одной оси радикала. [18]
На форму спектра ЭПР свободных радикалов существенно влияет взаимодействие их с матрицей, которое определяется природой твердой фазы и степенью ее кристалличности. От этого, в частности, зависит заторможенность движения радикалов или отдельных групп атомов радикала. На форму спектра ЭПР влияет и взаимодействие неспаренного электрона радикала с матрицей. Если при этом образуется одновременно несколько различных радикалов, то анализ спектров ЭПР часто становится очень сложной задачей. [19]
С, при которых находятся группы R. При движении верхнего радикала по окружности верхнего основания потенциальная энергия меняется не монотонно, а проходит через максимумы и минимумы. [20]
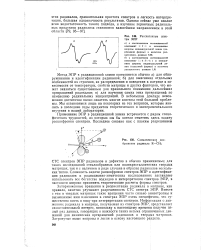 |
Рассчитанные спектры ЭПР.| Схематическое изображение радикала R - СН2. [21] |
Заторможенные вращения и реориентации радикала в матрице, как правило, заметно улучшают разрешенность СТС спектра ЭПР. Вместе с тем в твердых матрицах такие вращения часто сильно анизотропны и вызываемые ими изменения в спектрах ЭПР очень специфичны, что существенно иметь в виду при интерпретации спектров. Информация о движениях радикала в матрице, получаемая из спектров ЭПР, представляет самостоятельный интерес, поскольку к настоящему времени получен целый ряд данных, говорящих о важности таких весьма ограниченных движений для химических превращений радикалов в твердых матрицах. [22]
Форма линий зависит от характера движения радикалов, так как она определяется процессами релаксации. Различают два типа релаксации: спин-решеточную и спин-спиновую. Первая возникает из-за движения радикала, приводящего к флуктуациям магнитного поля, которое действует на неспаренный электрон. Если эти флуктуации имеют компоненту, осциллирующую с частотой, равной частоте перехода, то из-за этого может возникнуть переход. Время жизни верхнего состояния уменьшается, и в соответствии с принципом неопределенности энергетический уровень этого состояния уширяется. [23]
Проявляется также подвижность дипольно-группового типа, обусловленная радикалами, еще более удаленными от главной цепи. Такими кинетическими единицами являются концевые группы достаточно длинных боковых привесков. Так, в поли-ос-хлорметилметакрилатах наблюдается движение радикала, образованного концевой полярной группой, и его подвижность возрастает по мере удлинения алифатической части боковой цепи. [24]
На рис. 141 показано, при каких условиях происходит воспламенение смеси. При низких р и Т реакция не идет; увеличивая давление ( или температуру), мож-яо достичь нижнего предела воспламенения. Он объясняется тем, что с увеличением давления замедляется скорость движения радикалов к станкам сосуда и, следовательно, уменьшается вероятность их обрыва. Кроме нижнего для большинства взрывных процессов существует второй ( его называют варяним) лрещел воспламенения, связанный / с гибелью радикалов, в объеме при повышенных давлениях. В некоторых случаях ( как на рис. 141) наблюдается еще третий предел. Он, как правило, обусловлен тепловым взрывам. [25]
Заселенность s - орбиталей атомов А и В может возникать вследствие косвенных эффектов и обычно бывает небольшой. А и В 1с2 ( рхА) и с2 ( рхВ) ], вычисленная при помощи способа, описанного во второй главе, должна равняться единице. Значительные отклонения спиновой плотности от единицы могут быть связаны с такими причинами, как либрационное движение радикала около оси z или участие cf - орбиталей. Обе эти причины будут приводить к более низким значениям полной спиновой плотности. [26]
Сильное взаимодействие 1 и 2 с мицеллами ясно следует уже из того, что их растворимость в мицеллярных растворах в 100 раз превышает растворимость в воде. Наличие уширения резонасных линий и его величина также указывают, что 1 и 2 в значительной степени ассоциированы с мицеллами ДДС. Спектр ЭПР радикала 1 в воде состоит из трех линий одинаковой ширины, характерных для быстрого вращательного движения радикала. В 5 % - ном растворе ДДС линии ДДС заметно и притом неодинаково уширяются, что связано с торможением движения радикала вследствие его взаимодействия с мицеллой. [27]
 |
Времена корреляции тс зонда ТЕМПОЛ и спин-меченых полимеров в растворе и в клеточных суспензиях. [28] |
В последние годы значительные успехи в понимании этих вопросов связаны с использованием метода электронного парамагнитного резонанса ( ЭПР), позволяющего без нарушения нативности клеток слепить за молекулярной динамикой макромолекул вблизи поверхности и делать выводы о строении адсорбированного слоя. В ранее выполненных исследованиях [129, 130] методом ЭПР была изучена адсорбция флокулянтов на поверхности клеток E. В качестве полимеров, отличающихся по энергетике межмолекулярного взаимодействия с функциональными группами клеточной поверхности, были выбраны неионо-генный декстран и катионные полиэтиленимин и хитозан. Было установлено, что спектры ЭПР спин-меченого декстрана, слабо адсорбирующегося клетками, в супернатанте и суспензии клеток мало отличались друг от друга. Компонента замедленного движения принадлежит радикалам сегментов цепи, находящихся на границе клеточной стенки бактерии. Оцененное время тс трансляционно-поворотного движения радикалов ПЭИ включает вклады от крупномасштабного движения макромолекулярного клубка в целом и мелкомасштабного сегментального движения. [29]
Страницы: 1 2