Окислительное декар-боксилирование
Cтраница 1
Окислительное декар-боксилирование [56] ароматических кислот в присутствии солей меди ( при 200 - 350 С) дает фенолы или их эфиры в зависимости от используемого растворителя. Реакция используется для промышленного получения фенола из толуола через бензойную кислоту. Основные медные соли толуиловой и бензойной кислот при 200 - 220 С дают соответствующие о-гидроксикислоты, которые при более высоких температурах быстро декарбоксилируются. [1]
Кислоты, образующиеся в процессе окисления циклогевса-на, претерпевают два типа изменений - окислительное декар-боксилирование и этерификацию. [2]
Основным источником выделяемых при брожении газообразных продуктов ( С02 и Н2) служит реакция окислительного декар-боксилирования пирувата. У клостридиев описаны и другие пути образования молекулярного водорода. [3]
В каких биохимических процессах участвует коферментное производное тиамина: а) гликолиз; б) окислительное декар-боксилирование сс-кетокислот; в) окислительные реакции пентозофосфатного пути; г) неокислительные реакции пен-тозофосфатного пути; д) трансаминирование. [4]
Метилмасляная кислота, 1 - С14 - изомасляная кислота и 1 - С14 - изовалериановая кислота ( продукты окислительного декар-боксилирования соответственно изолейцина, валина и лейцина) превращаются в С14О2 в условиях, описанных для [ 3-окисления жирных кислот митохондриями арахиса ( см. стр. [5]
Все эти наблюдения, а также тот факт, что в отходящем газе кроме углекислоты обнаружена окись углерода до 1 %, позволяют предположить, что механизм окислительного декар-боксилирования уксусной кислоты достаточно сложен и, возможно, приводит к образованию целого ряда продуктов, что определяет целесообразность дальнейшего его исследования. [6]
 |
The Krebs Bicycle. ( Печатается с любезного разрешения д-ра David L. Nelson и д-ра М.М. Сох, 1993. [7] |
Следует, однако, подчеркнуть, что основным путем превращения углеводов в жиры является путь образования высших жирных кислот из ацетил - КоА, который образуется при окислительном декар-боксилировании пирувата. Последняя реакция практически необратима, поэтому образования углеводов из высших жирных кислот почти не происходит. Таким образом, синтез углеводов из жиров в принципе может происходить только из глицерина, хотя в обычных условиях реакция протекает в обратную сторону, т.е. в сторону синтеза жиров из глицерина, образующегося при окислении углеводов. Ацетил-Ко А, образующийся в процессе обмена углеводов, жиров и ряда аминокислот, служит пусковым субстратом как для синтеза жирных кислот ( а следовательно, и липидов вообще), так и для цикла трикарбоновых кислот. Для окисления ацетил - КоА в этом цикле требуется оксалоацетат, который является вторым ключевым субстратом в цикле Кребса. Оксалоацетат может синтезироваться из пировиноградной кислоты и СО2 благодаря реакции карбокси-лирования или образоваться из аспарагиновой кислоты в процессе транс-аминирования с а-кетоглутаратом. Следует подчеркнуть, что ацетоуксусная и 3-оксимасляная кислоты часто рассматриваются как транспортные формы активной уксусной кислоты, доставляющие ее для окисления в цикле Кребса в периферических тканях. Эти же реакции конденсации двух молекул ацетил - Ко А составляют начальные этапы синтеза холестерина, в свою очередь являющегося предшественником гормонов стероидной природы, витамина D3, а также желчных кислот. Последние в виде парных желчных кислот выполняют важную функцию эмульгаторов при переваривании липидов пищи в кишечнике, а также функцию транспортеров, способствуя всасыванию высших жирных кислот. Следует указать также на использование галактозы и частично глюкозы для биосинтеза цереброзидов и гликолипидов, выполняющих важные и специфические функции в деятельности ЦНС. В этом синтезе участвуют не свободные моносахариды, а гексозамины ( галактозамин и глюкозамин), биосинтез которых в свою очередь требует доставки амидного азота глутамина, интегрируя тем самым обмен углеводов, липидов и белков. [8]
Тем не менее специфические ферменты ( лактоназы) были идентифицированы в растениях и у многих видов бактерий. Эта реакция представляет собой обратимую реакцию окислительного декар-боксилирования, аналогичную реакциям, катализируемым НАДФ-зави-симой малатдегидрогеназой ( см. стр. В качестве промежуточного продукта в реакции участвует, вероятно, 3-кето-гексоновая кислота. [9]
Главным интермедиатом этой стадии является фосфонпируват ( РпРу), из которого могут быть получены природные ФОС с трехугольной цепочкой с использованием стандартных биосинтетических реакций, модифицирующих функциональные группы, включая и образование пептидов. Де-карбоксилирование РпРу образует ин-термедиаты с двухуглеродной цепочкой у фосфора, а окислительное декар-боксилирование одного из последних интермедиатов ( альдегида) приводит к одноуглеродному фосфонату. В свою очередь, одноуглеродный фосфонат ( фосфонмуравьиная кислота, PF) образует фосфонпируват ( скорее всего, реакцией переэтерификации), изомеризация которого, аналогичная изомеризации типа фосфат-фосфонат ( фос-фонат-фосфинат), приводит к соединению с двумя Р - С связями ( фосфи-наты) - путь, ведущий к образованию биалафоса. Образование природных ФОС с четырехуглеродной цепочкой у атома фосфора осуществляется действием ацетил-кофермента А по карбонильной группе соединения РпРу с последующим декарбоксилированием. [10]
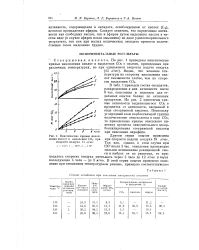 |
Состав оксидатов при окислении стеариновой кислоты.| Кинетические кривые накопления кислот и выделения СОа при скорости воздуха 12 л / час. [11] |
В табл. 1 приведен состав оксидатов, распределение в них активности после 8 час. На рис. 2 приведены кривые активности выделившегося С02 в процентах от активности, введенной в виде стеариновой кислоты. Поскольку углеродный атом карбоксильной группы количественно отщепляется в виде ССЬ, то приведенные кривые описывают кинетику процесса окислительного декар-боксилирования стеариновой кислоты при окислении парафина. [12]
Второй путь биосинтеза бензольного кольца в растениях носит название шикиматного ( схема 70, поз. Этот термин происходит от названия шикимовой кислоты 3.7 - одного из промежуточных звеньев биосинтетической последовательности. Она начинается реакцией продукта первичного метаболизма фосфоеяолпировиноградной кислоты с 3-фосфатом моносахарида эритрозы 3.3. В результате этого образуется 5-дегидрохинная кислота 3.5, которая через стадию хинной кислоты 3.6 превращается в кислоту ши-кимовую 3.7. Последняя является непосредственным предшественником таких производных бензола, как галловая 3.8 и протокатеховая 3.9 кислоты. Кроме того, от кислоты 3.7 проложен еще один важный путь биосинтеза. Он проходит через ее енольный эфир 3.10 с пировиноградной кислотой и хоризмовую кислоту 3.11. Последняя претерпевает кляйзеновскую перегруппировку в кислоту префеновую 3.12, дающую после окислительного декар-боксилирования я-гидроксифенилпировиноградную кислоту 3.13, которая служит биогенетическим предшественником фенилпропаноидов. Так называют метаболиты, в которых к бензольному кольцу присоединена прямая трех-углеродная цепь. [13]
Страницы: 1