Деление - клетка
Cтраница 3
При делении клетки двойные спирали расплетаются и, переходя к потомству, передают ему наследственные черты кодом последовательности молекулярных радикалов. [31]
При делении клетки цель разнимается посредине на две нитки, каждая переходит в свою клетку и там набирает себе дополнение. [32]
При делении клетки двойные спирали расплетаются и, переходя к потомству, передают ему наследственные черты кодом последовательности молекулярных радикалов. [33]
При делении клетки стебелек дихотомически ветвится. [34]
При делении клетки образуются две дочерние клетки, каждая из которых содержит хромосомы, ДНК которых являются фактически точными копиями молекулы ( молекул) родительской ДНК. Это простейший наблюдаемый процесс репликации. На практике, по одной из цепей родительского дуплекса оказывается в каждой дочерней клетке наряду с еще одной вновь синтезируемой комплиментарной цепью. Ферментом, который осуществляет этот новый синтез, является ДНК-полимераза III, которая подбирает подходящие дезоксинуклеотиды, в виде их 5 -трифосфатов, по принципу Уотсон-Криковского спаривания оснований к обеим раскрученным родительским цепям. [35]
При непрямом делении клеток имеет место удвоение генетического материала. [36]
В делении клетки тека не участвует. Обычно возникающие внутри ее - дочерние клетки ( или только одна из них) выходят наружу и образуют новую теку. [37]
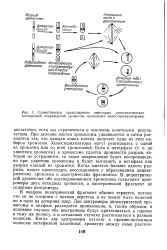 |
Схематическое представление некоторых цитогенетически. х последствий повреждений хромосом, вызванных хемостерилизаторами. [38] |
При делении клетки хромосомы удваиваются и затем расходятся так, что каждая новая клетка получает один из этих наборов хромосом. Хемостерилизаторы могут реагировать с одной из хроматид или со всей хромосомой. Когда имеется больше одного разрыва, может происходить воссоединение с образованием дицент-рических хромосом и ацентрических фрагментов. В дицентрическои хромосоме обе воссоединившиеся хромосомы сохраняют центромеры двух исходных хромосом, а ацентрический фрагмент не содержит центромеры. [39]
При делении клеток камбия число образующихся клеток ксилемы намного превышает число клеток флоэмы, причем у лиственных деревьев деление в сторону ксилемы происходит чаще, чем у хвойных. [40]
При делении клеток камбия, наряду с образованием клеток ксилемы, появляются клетки луба ( флоэмы), которые, подобно клеткам ксилемы, дифференцируются для выполнения различных функций. [41]
При делении материнской меристемной клетки возникают две дочерних клетки, одна из которых сохраняет способность к митозу и остается меристемной клеткой. Способные к делению клетки меристемы называют инициальными клетками, или инициалями. Образовавшиеся новые клетки называют производными клетка-м и. Некоторые из них могут делиться дальше, а другие дифференцируются ( специализируются), созревают и входят в состав тех или иных тканей, увеличивая их объем. Различают верхушечные ( апикальные) и боковые ( латеральные) меристемы. Первые находятся на концах главного и боковых побегов и в кончиках корней; они рассматриваются как первичные меристемы. Вторые располагаются параллельно боковым поверхностям ствола, побегов, корней и могут быть первичными и вторичными. [42]

Перед спорообразованием деление клеток прекращается, они резко увеличиваются в размерах. [44]
Карбин нарушает деление клеток у овса, овсюга, пшеницы. При действии его на растения наблюдаются ненормальности роста меристемы, прекращается развитие пазушных почек. Подавление развития пазушных почек является наиболее существенным в действии карбина. У пшеницы это действие выражается слабее и признаки его со временем проходят. [45]
Страницы: 1 2 3 4