Апикофильность
Cтраница 1
Апикофильность лиганда зависит от р - - взаимодействия, я-0 - взаимодействия, поляризуемости, пространственных факторов, а также от электроотрицательности ( см. разд. Апикофильность всегда вычисляется по отношению к другому лиганду, поскольку при перемещении одного лиганда из экваториального положения в апикальное, другой лиганд вынужден перемещаться в обратном направлении. [1]
Апикофильность лиганда зависит от р - - взаимодействия, я-а-взаимодействия, поляризуемости, пространственных факторов, а также от электроотрицательности ( см. разд. Апикофильность всегда вычисляется по отношению к другому лиганду, поскольку при перемещении одного лиганда из экваториального положения в апикальное, другой лиганд вынужден перемещаться в обратном направлении. [2]
Наиболее богатым источником информации об относительной апикофильности является динамическая спектроскопия ЯМР стабильных фосфоранов, данные которой интерпретируются с точки зрения изомеризации, включающей взаимный обмен двух апикальных заместителей на любые экваториальные. ЯМР I9F при различных температурах, осуществляются через промежуточные топомеры ( 25) и ( 26) ( схема 7), обладающие высокой энергией. При фиксированной группе В, энергия этого процесса изменяется симбатно с изменением апикофильности А по отношению к В. Свободная энергия активации - будет превышать разницу в апикофильности А и В на незначительную величину, вероятно, порядка 15 кДж / моль. [3]
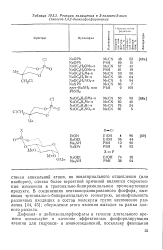 |
Реакции замещения в 2-галоген - 2-оксо ( тиоксо-1 3 2-диоксафосфоринанах. [4] |
В соединениях пентакоординированного фосфора, имеющих тригоналыю-бипирамидальную геометрию, апикофильность различных входящих в состав молекулы групп несомненно различна [44, 45]; обсуждение этого явления выходит за рамки данного раздела. [5]
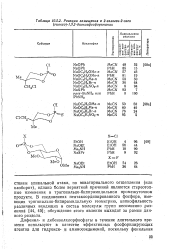 |
Реакции замещения в 2-галоген - 2-оксо ( тиоксо-1 3 2-диоксафосфоринанах. [6] |
В соединениях пентакоординированного фосфора, имеющих тригонально-бипирамидальную геометрию, апикофильность различных входящих в состав молекулы групп несомненно различий [44, 45]; обсуждение этого явления выходит за рамки данного раздела. [7]
Замещение будет сопровождаться сохранением конфигурации фосфора, если X обладает значительно более высокой апикофильностью, чем Y [ как, например, в ( 88) ], поскольку в этом случае псевдовращение будет преимущественно приводить к ( 95) с последующим отрывом уходящей группы из апикального положения. В системах, имеющих близкие по свойствам экзоциклические лиганды, связанные с фосфором, конечный состав продуктов будет зависеть от относительного времени жизни различных интермедиатов и способности групп X и Y к отщеплению. [8]
Апикофильность лиганда зависит от р - - взаимодействия, я-а-взаимодействия, поляризуемости, пространственных факторов, а также от электроотрицательности ( см. разд. Апикофильность всегда вычисляется по отношению к другому лиганду, поскольку при перемещении одного лиганда из экваториального положения в апикальное, другой лиганд вынужден перемещаться в обратном направлении. [9]
Апикофильность лиганда зависит от р - - взаимодействия, я-0 - взаимодействия, поляризуемости, пространственных факторов, а также от электроотрицательности ( см. разд. Апикофильность всегда вычисляется по отношению к другому лиганду, поскольку при перемещении одного лиганда из экваториального положения в апикальное, другой лиганд вынужден перемещаться в обратном направлении. [10]
Для понимания роли первоначально образующихся ТБП и дальнейшей изомеризации этих интермедиатов в процессе нуклео-фильного замещения у атома фосфора необходимо иметь возможность оценивать относительную стабильность различных, способных образовываться ТБП. Разница в энергии между двумя изомерными ТБП может быть рассчитана на основании изменения относительной способности лиганда занимать апикальное положение ( апикофильность) и изменения напряжения цикла в том случае, когда малый цикл перемещается из апикально-экватори-ального ( ае) в диэкваториальное ( ее) положение. [11]
Для понимания роли первоначально образующихся ТБП и дальнейшей изомеризации этих интермедиатов в процессе нуклео-фильного замещения у атома фосфора необходимо иметь возможность оценивать относительную стабильность различных, способных образовываться ТБП. Разница в энергии между двумя изомерными ТБП может быть рассчитана на основании изменения относительной способности лиганда занимать апикальное положение ( апикофильность) и изменения напряжения цикла в том случае, когда малый цикл перемещается из апикально-экваторн-ального ( ае) в диэкваториальное ( ее) положение. [12]
Наиболее богатым источником информации об относительной аппкофильности является динамическая спектроскопия ЯМР стабильных фосфоранов, данные которой интерпретируются с точки зрения изомеризации, включающей взаимный обмен двух апикальных заместителей на любые экваториальные. Изменения в окружении обеих СР3 - групп, фиксируемые с помощью спектроскопии ЯМР 19F при различных температурах, осуществляются через промежуточные топомеры ( 25) и ( 26) ( схема 7), обладающие высокой энергией. При фиксированной группе В, энергия этого процесса изменяется симбатно с изменением апикофильности А по отношению к В. [13]
Наиболее богатым источником информации об относительной апикофильности является динамическая спектроскопия ЯМР стабильных фосфоранов, данные которой интерпретируются с точки зрения изомеризации, включающей взаимный обмен двух апикальных заместителей на любые экваториальные. ЯМР I9F при различных температурах, осуществляются через промежуточные топомеры ( 25) и ( 26) ( схема 7), обладающие высокой энергией. При фиксированной группе В, энергия этого процесса изменяется симбатно с изменением апикофильности А по отношению к В. Свободная энергия активации - будет превышать разницу в апикофильности А и В на незначительную величину, вероятно, порядка 15 кДж / моль. [14]
Из рассмотренных реакций видно, что состав продуктов щелочного гидролиза фосфониевых солей определяется сочетанием различных факторов. Если атом фосфора не входит в состав малого цикла, гидролиз ацилфосфониевых солей осуществляется путем нуклеофильной атаки на углерод карбонильной группы, однако, если фосфор является частью малого цикла, то нуклео-фильная атака направлена на атом фосфора вследствие ослабления напряжения в цикле. Когда напряжение цикла велико, например в фосфолах или фосфетанах, происходит расширение цикла; при меньшем напряжении промежуточный продукт имеет большее время жизни, достаточное для перемещения обладающей высокой апикофильностью ацильной группы в апикальное положение и последующего ее отщепления. [15]
Страницы: 1