Длина - молекула
Cтраница 2
Длина молекул эфира себациновой кислоты, равная 25 А, по меньшей мере вдвое больше длины молекулы эфира тиодигликолевой кислоты. К тому же в составе последнего эфира имеется атом серы, что увеличивает ван-двр-ваальсовское взаимодействие с макромолекулами. Несмотря на эти благоприятные факторы в данном случае, очевидно, не происходит образования сольватных оболочек. В других опытах было показано, что эфиры тиодигликолевой кислоты до 200 С не растворяют триацетат целлюлозы, хотя их можно вводить в пленку в больших количествах. [16]
Длина молекулы ДНК хромосомы человека достигает 8 см, но умещается в хромосоме длиной в несколько нанометров. Это объясняется тем, что двухцепочечная спираль ДНК в пространстве укладывается в еще более сложную кольцевую форму, или суперспираль. [17]
Длина молекулы ДНК хромосомы человека достигает 8 см, но умещается в хромосоме длиной в несколько нанометров. [18]
Увеличение длины молекулы до 1500 - 2000 звеньев дает твердый и гибкий пластик, из которого можно делать гибкие пленки и трубы. Полиэтилен с молекулой большой длины ( 5 - 6 тыс. звеньев) обладает более высокой твердостью и применяется для изготовления жестких труб, контейнеров. [19]
Изменения длины молекул субстрата и фермента равны х и у соответственно. [20]
Увеличение длины молекулы диамина приводит к повышению гибкости цепей, что, видимо, облегчает релаксацию внутренний напряжений. В результате прочность пленок увеличивается. [21]
Например, длина молекулы этилена составляет 0 133 нм, а высокомолекулярного соединения полиэтилена - 1000 - 10 000 нм. [22]
А меньше длины молекулы тетрагидрофурфурилолеата, растворяющая способность вследствие отсутствия гетероциклического ядра ослаблена и до 190 С наблюдается только частичное растворение поливинилхлорида. Снижение растворяющей способности бутилового эфира рицин-олеиновой кислоты по сравнению с тетрагидрофурфурилолеатом объясняется тдкже изменением полярности молекулы вследствие наличия в ней ОН-групп. [23]
Для измерения длины молекул выбирают обособленные, по возможности, вытянутые, целые и неповрежденные молекулы ДНК. Длина молекулы определяется в ангстремах делением длины изображения нити ДНК на конечное увеличение снимка. Конечное увеличение представляет собой произведение первичного увеличения, получаемого на негативе, и увеличения при фоторепродукции. Значения первичного увеличения для всех ступеней электронного микроскопа должны быть точно програ-дуированы по диффракционной решетке и глобулам латекоа. [24]
С увеличением длины молекулы расход тепла на плавление повышается. [25]
При увеличении длины молекулы парафина площадь свободных зон уменьшится, что приведет к возрастанию плотности взаимодействующих с поверхностью водородных и углеводо ролных атомов и к увеличению энергии взаимодействия слон с поверхностью в целом, а следовательно, п силы сдвига. [26]
Прямые измерения длины молекул ДНК из этих организмов, проведенные методом радиоавтографии и электронйой микроскопии ( фиг. [27]
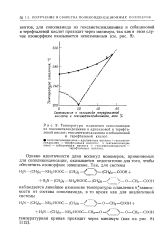 |
Температура плавления сополиамидов. [28] |
Однако идентичности длин молекул мономеров, применяемых для сополиконденсации, оказывается недостаточно для того, чтобы обеспечить изоморфное замещение. [29]
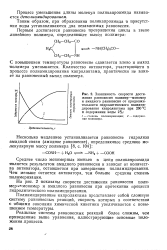 |
Зависимость скорости достижения равновесия полимер-мономер и амидного равновесия от продолжительности гидролитического полиамидирования капролактама при 260 С и содержании воды 2 %. [30] |
Страницы: 1 2 3 4