Катион - аминокислота
Cтраница 1
Катионы аминокислот замещают катионы буфера и таким образом связываются со смолой. Затем через колонку пропускают буфер, который содержит катионы, вытесняющие аминокислоты, после чего, меняя концентрацию буфера, элюируют ( вымывают) индивидуальные аминокислоты в очередности, которую можно предсказать заранее. После элюирования аминокислота реагирует с нингидрином. [1]
Можно предположить, что различные катионы, в частности катионы аминокислот, образуют солеобразные соединения с сульфогруппами полимерного комплекса. [2]
 |
Зависимость положения максимума сорбции ( pHj от.| Зависимость положения максимума сорбции ( рН от. [3] |
Механизм сорбции аминокислот Н - формой сульфокатионита заключается в сорбции катиона аминокислоты, причем выделяющиеся в раствор ионы водорода сдвигают равновесие цвиттерион - катион в сторону образования катиона, который и сорбируется. В случае же сорбции аминокислот Na-формой смолы выходящие в раствор ионы Na препятствуют дальнейшей сорбции аминокислоты из-за обычной конкуренции Na - HOHOB; кроме того, происходит защелачивание раствора, уменьшается концентрация катионов аминокислоты, а следовательно, и сорбированное количество аминокислоты. [4]
Согласно уравнениям реакций ( 19) и ( 20) в очень кислых средах в растворе будут преобладать катионы аминокислоты, в сильнощелочных средах аминокислота будет в форме аниона. [5]
Один из способов преодоления трудностей в количественном изучении ионообменной сорбции аминокислот состоит во введении диссоциационных представлений, например в предположении о сорбции наряду с катионами аминокислот их диполярных ионов, что, естественно, должно зависеть от рН и не только внешнего раствора, но и гелевой фазы ионйта. Второй вариант подхода заключается в использовании представления об изменчивости коэффициента активности резинатов при изменении рН внешнего ( и внутреннего) раствора, причем эти изменения могут быть существенными. [6]
Термодинамический анализ сорбции моноаминомонокарбоно-вых аминокислот ( и аналогичных им пептидов в электрохимическом отношении) в нейтральной области рН для сульфокатионитов в водородной форме может быть осуществлен с использованием различных молекулярных форм компонентов в сосуществующих фазах: диполярного иона в растворе и резината, включающего катион аминокислоты в фазе ионйта, а также резината в водородной форме. [7]
 |
Зависимость положения максимума сорбции ( pHj от.| Зависимость положения максимума сорбции ( рН от. [8] |
Механизм сорбции аминокислот Н - формой сульфокатионита заключается в сорбции катиона аминокислоты, причем выделяющиеся в раствор ионы водорода сдвигают равновесие цвиттерион - катион в сторону образования катиона, который и сорбируется. В случае же сорбции аминокислот Na-формой смолы выходящие в раствор ионы Na препятствуют дальнейшей сорбции аминокислоты из-за обычной конкуренции Na - HOHOB; кроме того, происходит защелачивание раствора, уменьшается концентрация катионов аминокислоты, а следовательно, и сорбированное количество аминокислоты. [9]
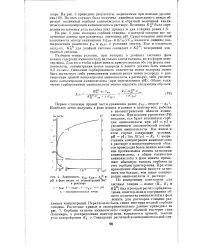 |
Зависимость g рН в фазе смолы от концентрации Na в растворе. [10] |
Мы имеем в этом случае следующие условия: рН pJ; lim / ( Сн) - i - С возрастанием концентрации аминокислоты в растворе в изоэлектрической области происходит более полное заполнение противоионов ионита катионами аминокислоты, а общее количество аминокислоты в фазе ионита превышает обменную емкость сорбента за счет сорбции цвиттер-иона. [11]
Число исследований, посвященных изучению механизма сорбции аминокислот ионитами, сравнительно невелико, а взгляды, высказываемые отдельными авторами, различны и порой противоречивы. Картен и Каннан [1], изучавшие сорбцию нейтральных аминокислот на сильнокислом катио-ните из растворов аминокислот с различным значением рН, предполагают, что при рН, близких к pj, происходит либо обычный ионный обмен противо-ионов водорода с катионами аминокислоты, присутствующими в малой концентрации, либо идет непосредственное присоединение цвиттер-иона с переходом иона водорода к карбоксильной группе аминокислоты. [12]
Белки представляют собой амфотерные соединения, моделируемые аминокислотами в электрохимическом отношении. Однако положение изоэлектрической точки для них может варьироваться в широких масштабах. Возвращаясь к описанному выше механизму сорбции диполярных ионов и существованию резинатов моноаминомонокарбоновых кислот в состоянии, включающем катионы аминокислот, естественно выдвинуть предположение о том, что подобный эффект зависит от расстояния между аминными и карбоксильными группами и что для белков он не должен всегда выполняться. Следует отметить, что состояние резинатов белков до настоящего времени детально не обследовалось, в связи с чем предлагаемые модели для сорбции белков ионитами следует рассматривать с определенной долей осторожности. [13]
Сено и Ямабе [9, 10] дают опытные кривые зависимости величины сорбированного количества аминокислот ионитами от рН раствора и приводят метод расчета концентрации катионной формы аминокислоты в растворе. Они считают, что сорбция аминокислот подчиняется законам ионного обмена. Достоинством их работы является то, что при определении константы ионного обмена аминокислота - водород они предлагают оперировать не аналитической концентрацией, а концентрацией катионов аминокислоты в растворе. [14]
Было показано, что сорбция аминокислот в подобных системах может быть описана при помощи обычного уравнения ионного обмена, однако для расчетов следует использовать концентрации катионов аминокислоты в растворе и в ионите. [15]
Страницы: 1