Кинетика - рост
Cтраница 1
Кинетика роста и легирования слоев в зависимости от температуры кристаллизации исследовалась на слоях ориентации 2 от ( 111) А, которые выращивались в системе GaAs-HI - На. [1]
 |
Константы скорости роста цепи при полимеризации стирола. [2] |
Кинетика роста цепи в большой мере определяется эффективной константой скорости роста. В табл. 1.6 приведены значения констант роста для различных механизмов полимеризации стирола. [3]
Кинетика роста сопротивления фильтротканей в значительной степени определяется структурными особенностями засорений, закрепляющихся на волокнах тканей. [4]
Кинетика роста пленок зависит от стадий их формирования. На начальной стадии окисления образование первых слоев окисла происходит таким образом, чтобы решетка оксида приблизилась по своим параметрам к решетке исходного металла. На чистой поверхности металла образование первичной оксидной пленки происходит так, чтобы решетка оксида приближалась по своим параметрам к решетке исходного металла. Внешняя часть слоя заряжена отрицательно, а пограничный с металлом слой положительно. [5]
Кинетика роста реальных кристаллов определяется, таким образом, структурой поверхности кристалла-подложки. [6]
Кинетика роста удельной активности и кинетика накопления гидроперекисей позволяют по формулам изотопного кинетического метода рассчитать раздельно скорости образования и расходования гидроперекисей в процессе окисления. [7]
 |
S. Распределение твердости в зоне сплавления среднелегированной. [8] |
Кинетика роста толщины прослоек в соответствии с закономерностями диффузионного процесса определяется экспоненциальной зависимостью от температуры и квадратичной от времени выдержки. В координатах log 6 ( 7) и 5 ( л / О рост толщины прослоек 5 отображается прямыми линиями. Результатом их развития является создание зон переменного состава, структуры и твердости. [9]
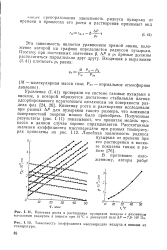
Кинетику роста и растворения исследовали при разных АР для пузырьков такого размера, что влиянием поверхностной энергии можно было пренебречь. Полученные результаты ( рис. 1.11) показывают, что принятая модель процесса в данном случае хорошо оправдывается. [11]
Кинетику роста трещин, протяженность которых сопоставима с размерами характерных элементов микроструктуры, необходимо рассматривать в микромасштабе. Развитие трещин в таком масштабе является дискретным и зависит от свойств отдельных фрагментов микроструктуры, а статистическая природа усталостного разрушения проявляется особенно ярко. В этих условиях понятия сплошности среды и ее однородности, а также аппарат механики сплошной среды могут оказаться неприемлемыми. [12]
Для кинетики роста ядер важное значение имеет кристаллическая структура поверхности раздела. Во-первых, поверхность раздела может быть полностью некогерентной в том смысле, что между атомами исходного вещества и атомами продукта может не существовать никакого кристаллографического соответствия. В этом случае взаимодействие между продуктом и реагирующим веществом определяется соотношением их объемов. Насколько известно автору, примерно такую форму имеют только ядра, образующиеся при дегидратации на некоторых из граней гептагидрата сульфата никеля и пентагид-рата сульфата меди. Но даже в этих случаях такая форма ядер может определяться диффузией, происходящей предпочтительно в некоторых из возможных направлений. [13]
Рассчитана кинетика роста твердой фазы и доменная структура бислоя при его двумерной кристаллизации Экспериментальные данные, полученные ультразвуковым методом, звидетельствуют о различиях кинетики гетерофазных флуктуации плотности при ионной, молекулярной, изотопной и структурной модификации ЛЖК. [14]
Уравнения кинетики роста, образования продуктов, потребления субстратов, автолиза биомассы и инактивации продуктов являются специфическими для микробиологических процессов. Математическое описание кинетики этих процессов отличается от традиционной химической кинетики, поскольку все процессы осуществляются с участием биокатализаторов-ферментов. Причем субстрат в процессе превращения в организованную биомассу или продукт метаболизма проходит весьма большое число промежуточных стадий биохимических ферментативных реакций. Поскольку большинство биохимических реакций осуществляется в клетках микроорганизмов, в микробиологической кинетике принято в качестве выходных параметров использовать не абсолютные значения скоростей реакций, а удельные, отнесенные к единице веса микробной массы. [15]
Страницы: 1 2 3 4 5