Абсцизовая кислота
Cтраница 3
Следует отметить, что природные ингибиторы - абсцизовая кислота и фенольные соединения - легко вымываются водой и могут входить в состав выделений, влияющих на прорастание семян и рост проростков растений в сообществах. [31]
В литературе имеются первые данные о действии абсцизовой кислоты на биосинтез гиббереллинов у высших растений. [32]
Эффект торможения был имитирован и с помощью экзогенно введенной абсцизовой кислоты, которая вызывала резкое торможение прорастания семян. [33]
К ан-титранспирантам метаболического типа некоторые авторы относят абсцизовую кислоту, алахлор, алкенилянтарную кислоту, хлормекват, 2-хлормеркур - 4 6-динитрофенол, даминозид, 2 4 - динитрофенол, 8-оксихинолин, ИУК, хлорфлюренол, фенил-меркурацетат, салицил-альдоксим. [34]
В данной работе предлагается метод выделения и идентификации абсцизовой кислоты из почек черной смородины, но он не пригоден для анализа и других растительных объектов. [35]
Дается краткий литературный обзор, касающийся выделения и определения абсцизовой кислоты в растениях. Приводится оригинальный метод выделения и идентификации абсцизовой кислоты из почек черной смородины. [36]
За последнее десятилетие опубликовано много статей, посвященных влиянию абсцизовой кислоты на кор-необразование, однако приводимые результаты противоречивы. Следует отметить, что многие из этих исследований проводили для выяснения роли регуляторов роста в индукции корнеобразования. [37]
Если к хроматину, обладавшему РНК-полимеразной активностью, добавляли абсцизовую кислоту ( 10 - 6 М), то это приводило к торможению полимеразной активности на 22 - 38 % в том случае, когда этот ингибитор вводили в систему гомогенизированных проростков. Эти данные Пирсон и Вэринг интерпретируют как возможность абсцизовой кислоты влиять на РНК-полимеразную активность только после связывания этого ингибитора с эндогенным цитоплазмати-ческим фактором. [38]
Тильберга ( Tillberg, 1970) было показано, что абсцизовая кислота не влияет на процесс выделения кислорода и фото-фосфорилирования. Автор считает, что в комплексе Р - ингибитора присутствуют другие вещества, может быть фенолы, которые оказывают на эти процессы тормозящее действие. Хотя данных по тормозящему действию фенолов на окислительное и фотофосфорилиро-вание имеется довольно много, все-таки нельзя признать эту проблему хорошо разработанной. [39]
Если фенолы действуют главным образом на активность ферментов, то абсцизовая кислота в основном подавляет их синтез. [40]
Возникает вопрос о поведении в толуоле нового ингибитора роста - абсцизовой кислоты. При введении 0 5 мкг абсцизовой кислоты в пятно вся ее ингибиторная активность ( 40 % роста контрольных отрезков колеоптилей) после разделения в толуоле оставалась на старте, однако опытов с абсцизовой кислотой в присутствии хлорофилла не проводилось. [41]
Таким образом, производные р-ионона - полупродукты биосинтеза () абсцизовой кислоты - обладали слабым ингибиторным действием ( С50 800 - 160 мг / л) на рост отрезков колеоптилей. [42]
Сравнение биологической активности ингибитора из почек черной смородины с биологической активностью абсцизовой кислоты, кумарина и салициловой кислоты показало, что ингибитор из почек черной смородины вызывает подавление роста, характерное для абсцизовой кислоты. [43]
Действие гетероауксина на живое растение тесно скоординировано с эффектами прочих фитогормонов: абсцизовой кислоты 2.164, гибберелли-нов ( разд. Искусственное добавление индолилуксусной кислоты нарушает их естественный баланс, и ответ растительного организма неоднозначен. Поэтому, хотя добавлением гормона можно добиться ускорения роста и увеличения плодов, практическое использование этого свойства в полевых условиях затруднено. Гетероауксин с успехом применяется при выращивании саженцев черенкованием, так как стимулирует образование корней и развитие корневой системы. [44]
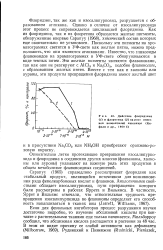 |
Действие флоридзина ( / и флоретина ( 2 на рост отрезков колеоптилей пшеницы ( Ке-фели и др., 1969 б. [45] |
Страницы: 1 2 3 4