Концентрация - аминокислота
Cтраница 2
 |
Образование 2 4-динитрофенильных производных амииокислетг.| Образование дипептида. [16] |
При надлежащим образом подобранных условиях интенсивность окраски можно использовать для колориметрического определения концентраций аминокислот, так как этот метод обладает очень высокой чувствительностью. [17]
Определив предварительно константы а ж Ь, можно по величине S определить концентрацию аминокислот в исходных растворах. [18]
Определив предварительно константы а и Ъ, можно по величине S определить концентрацию аминокислот в исходных растворах. [19]
Томпкинс предполагает, что эти аминокислоты, первоначально служили внутриклеточными символами, отражающими изменение концентрации аминокислот в среде, но в последующем стали использоваться для передачи сигналов на коротком расстоянии междуклетками нервной системы. [20]
Показатели бактериального выщелачивания золота зависят от содержания аминокислот в культуральных растворах: при увеличении концентрации аминокислот от 0 5 - 1 до 3 5 - 5 г / л концентрация золота в растворах увеличивается в 3 - 4 раза. Предпочтительны большие отношения Ж: Т и слабощелочная среда выщелачивающего раствора. Золото из получаемых бактериальных вы-щелатов успешно извлекается сорбцией на анионитах или активированном угле. [21]
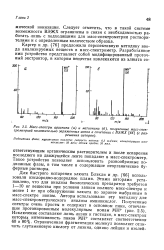 |
Масс-спектры аргинина ( а и метионина ( б, полученные масс-спек-трометрией положительно заряженных ионов в сочетании с ВЭЖХ ( с разрешения авторов. [22] |
Подвижная фаза: муравьиная кислота; скорость потока: 0 5 мл / мин; концентрация аминокислот: 50 мг. [23]
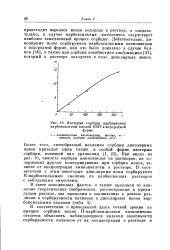 |
Изотерма сорбции альбомицина. [24] |
Как видно из рис. 16, емкость сорбции аминокислот из растворов, не содержащих других конкурирующих при сорбции ионов, зависит от концентрации аминокислоты в растворе. В соответствии с этим некоторые диполярные ионы сорбируются Н - карбоксильными смолами из разбавленных растворов с небольшими емкостями. [25]
Реакция комплексообразования аминокислоты ( гистидина) с ионами Ni2 позволяет по второй волне, соответствующей восстановлению аминокислотного комплекса двухвалентного Ni, определить концентрацию аминокислоты. Реакция образования ацетиленида ртути была использована для определения ацетилена. [26]
На полученных хроматограммах проводят визуальное сравнение пятен анализируемого и стандартных растворов по размеру и интенсивности окраски, на основании чего делают вывод о концентрации определенной аминокислоты. При наличии планиметра определяют концентрацию аминокислоты в анализируемом растворе, измеряя площади пятен. [27]
Величина оптической плотности раствора при анализе аминокислот зависит в сильной степени от условий хроматографирования ( качества бумаги, числа пропусканий растворителей и др.), поэтому для определения концентрации аминокислоты в анализируемом растворе нельзя пользоваться заранее сделанными калибровочными кривыми. Здесь пользуются установленной зависимостью между концентрацией кислоты и оптической плотностью раствора, которая в пределах концентраций аминокислот от 0 025 до 0 25 мкмоль ( микромолей) имеет линейный характер. [28]
Недавние исследования показали, что, хотя начальные изменения в клеточной осмоляльности определяются изменениями в концентрации неорганических ионов, осмотическая адаптация тканевых клеток в большей своей части бывает обусловлена сдвигами концентраций внутриклеточных аминокислот. У ракообразных главную роль в изменениях общей внутриклеточной осмоляльности играют глицин, аланин, пролин и глутаминовая кислота, хотя относительный вклад каждого из этих веществ у разных видов различен. Echino-dermata) в регуляции клеточной осмоляльности участвуют только две аминокислоты - глицин и таурин. У кольчатого червя Arenicola главную роль в адаптации клеток к осмотическим сдвигам играют глицин, аланин и глутаминовая кислота, тогда как концентрация пролина не изменяется. [29]
Поскольку между удельной радиоактивностью и концентрацией аминокислот существует линейная зависимость, то можно сказать, что удельную радиоактивность нефракционированного белка для образца, весящего 100 мкг, или для белковой зоны в микрогеле можно сравнивать с эквивалентными радиоактивностями образцов из идентичных участков мозга другого полушария того же животного, а также с образцами мозга других животных путем деления удельной радиоактивности белка на концентрацию аминокислот, содержащих тритий. Все значения удельной радиоактивности, таким образом, сравнивают при одной и той же концентрации аминокислоты, меченой тритием, названной здесь единичной концентрацией 3Н - лейцина. [30]
Страницы: 1 2 3 4