Отдельные аминокислота
Cтраница 3
Приведенные данные по поводу незаменимости отдельных аминокислот для роста или азотистого равновесия были первоначально получены в опытах на крысах и собаках. Имеющиеся в настоящее время данные позволяют предполагать, что для поддержания азотистого равновесия у людей необходимы все вышеуказанные незаменимые аминокислоты, за исключением, по-видимому, аргинина и гистидина. В аналогичных опытах на цыплятах выяснилось, что гликокол является аминокислотой, незаменимой для роста цыплят. Но эти данные отличаются от данных, полученных в опытах на собаках и крысах. Поэтому следует предостеречь от механического переноса результатов опыта с одного вида животных на другие. Кроме того, не следует забывать того важного обстоятельства, что заменимые аминокислоты существенно влияют на потребность в незаменимых аминокислотах. Потребность, например, в метионине определяется содержанием цистина в диете; чем больше в пище имеется цистина, тем меньше расходуется метионина для биологического синтеза цистина. Последний уменьшает, следовательно, потребность организма в метионине. Наконец, если в организме скорость синтеза какой-либо заменимой аминокислоты становится недостаточной, то появляется повышенная потребность в ней, которая может быть компенсирована поступлением ее с пищей. Отсюда ясна условность деления аминокислот на заменимые и незаменимые. [31]
Приведенные данные по поводу незаменимости отдельных аминокислот для роста или азотистого равновесия были первоначально получены в опытах на крысах и собаках. Имеющиеся в настоящее время данные позволяют предполагать, что для поддержания азотистого равновесия у людей необходимы все вышеуказанные незаменимые аминокислоты, за исключением, по-видимому, аргинина и гистидина. В аналогичных опытах на цыплятах выяснилось, что гликокол является аминокислотой, незаменимой для роста цыплят. Но эти данные отличаются от данных, полученных в опытах на собаках и крысах. Поэтому следует предостеречь от механического переноса результатов опыта с одного вида животных на другие. Кроме того, не следует забывать того важного обстоятельства, что заменимые аминокислоты существенно влияют на потребность в незаменимых аминокислотах, потребность, например, в метионине определяется содержанием цистина в диете; чем больше в пище имеется цистина, тем меньше расходуется метионина для биологического синтеза цистина. Последний уменьшает, следовательно, потребность организма в метионине. Наконец, если в организме скорость синтеза какой-либо заменимой аминокислоты становится недостаточной, то появляется повышенная потребность в ней, которая может быть компенсирована поступлением ее с пищей. Отсюда ясна условность деления аминокислот на заменимые и незаменимые. [32]
Белки синтезируются на рибосомах из отдельных аминокислот, образуемых самими микроорганизмами. Исключение составляют некоторые ауксотрофные мутанты, для которых необходимо присутствие в среде определенных аминокислот. Биосинтез аминокислот в клетке идет ферментативно из неорганического азота и различных соединений углерода, например продуктов аэробного или анаэробного разложения углеводов. [33]
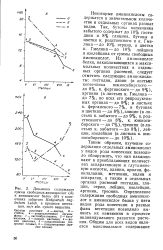 |
Динамика содержания суммы свободных аминокислот ( А и аминокислот белка ( Б у копеечника забытого Hedysarum neg-lectum Ledeb. в процессе вегетации, мг / г абс. сухого вещества. [34] |
Таким образом, изучение содержания отдельных аминокислот у видов рода копеечник позволило обнаружить, что они накапливают в преобладающих количествах аспарагиновую и глутамино-вую кислоты, аланин, пролин, фе-нилаланин, метионин, валин и аспарагин, а также в отдельных органах растений гистидин, глицин, серии, лейцин, изолейцин, аргинин, треонин. [35]
Такое протеолитическое расщепление белков до отдельных аминокислот имеет большое значение в питании животных. Белки, будучи коллоидами, не диффундируют через пористые перегородки ( не диализируют) и не могут сквозь стенки кишечника проникнуть в кровь; кристаллические аминокислоты, на которые белок распадается в результате гидролиза, хорошо растворимы в воде и легко диффундируют сквозь стенки кишечника. Из этих аминокислот животное строит в соответствующих органах свои белки. [36]
Получение белков, витаминов, отдельных аминокислот и других ценных веществ методами биосинтеза в настоящее время является весьма актуальной проблемой. В основе этого процесса лежит реакция окисления углеводородов в присутствии ферментативных систем. [37]
Отличия состояли в появлении на хромато-лектрофореграм-мах отдельных аминокислот, характерных для определенной расы, и нерасшифрованных пятен, расположенных неодинаково у различных рас фитофторы, а также в величине и интенсивности пятен идентифицированных аминокислот. [38]
Существует целый ряд других реакций на отдельные аминокислоты, которые так же используются для открытия белка. К ним относятся следующие реакции. [39]
Существует целый ряд других реакций на отдельные аминокислоты, которые также используются для открытия белка. К ним относятся следующие реакции. [40]
Некоторые виды Clostridium способны разлагать также отдельные аминокислоты, например серии, гистидин, глутамино-вую кислоту, метионин, тирозин; к числу продуктов распада относятся аммиак, водород и углекислота. [41]
Что касается механизма синтеза белка из отдельных аминокислот, то здесь наши сведения отличаются крайней недостаточностью. Было высказано предположение о возможности осуществления этого синтеза также под влиянием протеиназ. Эта точка зрения основывается на примерах обратимости ферментативного действия. Из нее следует, что катепсины могут не только гидролизовать белок на аминокислоты, но и ресинтезировать из них белок. [42]
Несмотря на интенсивное самообновление, соотношения отдельных аминокислот в белке являются довольно постоянными. [43]
В таблице 1 приведены результаты определения отдельных аминокислот в гидролизатах мощного чернозема Граковского-опытного поля НИУИФ, подзолистой суглинистой почвы Долгопрудной агрохимической опытной станции и той же почвы, компостировавшейся в течение двух лет с соломой и минеральным азотом. [44]
Несмотря на интенсивное самообновление, соотношение отдельных аминокислот в белке является довольно постоянным. Это в известной мере дает основание предполагать, что процессу обновления белка должно предшествовать образование всего набора аминокислот, составляющих белковую молекулу. [45]
Страницы: 1 2 3 4