Основная масса - белок
Cтраница 2
 |
Хроматографирование активной. [16] |
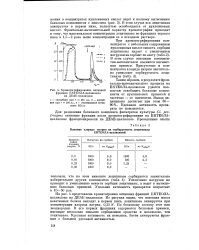
На рис. 4 представлена хроматограмма активных фракций ЕКТЕОЛА-целлюлозы на ДЕАЕ-целлюлозе. Из рисунка видно, что основная масса неактивных белков выходит в первом пике, лецитиназа элюируется при концентрации хлорида натрия, 0 25 N. По белковому составу этот пик неоднороден. В его первых фракциях содержатся белковые примеси, имеющие несколько меньшее сорбционное сродство к целлюлозе, чем лецитиназа. Удельная активность их невелика. Затем элюируются фракции с большей удельной активностью. [17]
Основной надосадочный раствор и промывную жидкость, полученную при отделении нерастворимого в воде геля, объединяют, концентрируют и лиофилизуют ( стр. В результате получают груп-поспецифическое вещество крови, свободное от основной массы посторонних белков и жиров. [18]
Метод основан на том, что гликоген хорошо растворим в воде и достаточно устойчив в слабокислой среде. Поэтому метод выделения гликогена сводится к механическому разрушению ткани и экстракции гликогена 5 % - ным раствором ТХУ. Основная масса белков при процедуре денатурирует и их легко удалить из раствора фильтрованием. [19]
Оставался еще неясным вопрос, не является ли кислотность белковых компонентов мембран и рибосом специфической особенностью именно этих субклеточных структур или же это фундаментальное свойство большинства белков галофильной клетки. По крайней мере частичный ответ на этот вопрос был получен в 1970 г. при сравнительном изучении основной массы белков у галофильных и близких к ним негалофильных бактерий. Выражение основная масса носит условный характер и означает белок, осаждаемый кислотой после удаления клеточных стенок и мембран. Эта белковая фракция составляет около 80 % всего белка клетки и включает большую часть растворимых ферментов. Как и следовало ожидать, эти белки отличаются необычайно высоким содержанием кислых аминокислот. Молярное отношение кислых аминокислот к основным достигает здесь примерно 10: 1, что даже выше соответствующих величин для белков рибосом и клеточных мембран. Таким образом, у крайне галофильных форм бактерий, по-видимому, большинство белков, если не все они, требуют высокой концентрации солей по той же причине: необходимо нейтрализовать избыток отрицательных зарядов, который в отсутствие катионов ведет к появлению сил электрического отталкивания и к нарушению нормальной структуры и функции. Поэтому в заключение нужно будет рассмот-реть влияние катионов на функции галофильных белков. [20]
Белки растительного происхождения получают главным образом из семян злаков и из бобовых растений. Семена, из которых предполагают выделить белки, должны быть хорошо измельчены и, если необходимо, обезжирены при помощи диэтилового или петролейного эфира. Извлечение белков из размельченного и обезжиренного семенного материала производится при помощи водных растворов солей, буферных растворов или этилового спирта. Основная масса белков, извлеченных этим путем из семян, относится к глобулинам и только от 0 1 до 0 5 % белков - к белкам типа альбумина. [21]
Данные исследований свидетельствуют о том, что природа и функция ферментативного аппарата, осуществляющего синтез белков в споре и материнской вегетативной клетке, одинаковы. Основной системой транспорта электронов в спорах является растворимая флавопротеино-ксидаза. При споруляции выявлена высокая активность ряда ферментов, например аргиназы, некоторых нуклеаз. Основная масса белков споры синтезируется заново в период споруляции. Одновременно отмечается процесс распада белков, причем многими авторами обнаружена тесная взаимосвязь протеолитиче-ской активности с функцией спорообразования у бактерий. Из вновь синтезированных белков лишь незначительная часть используется для создания структурных элементов споры. [22]
Проламины выделяют по следующей методике. Спиртовой экстракт из мерной колбы переливают в прибор для отгонки спирта в вакууме. После полного удаления спирта к оставшемуся раствору, в котором белки начинают выпадать в осадок, добавляют небольшое количество воды и оставляют в холодильнике в течение ночи. При этом основная масса белков выпадает в осадок. Содержимое колбы переносят в центрифужные пробирки, колбы споласкивают ацетоном и осадок белков отделяют центрифугированием при 3 - 4 тыс. об / мин, в течение 5 - 10 минут. Затем белки в пробирках несколько раз промывают ацетоном и эфиром с последующим центрифугированием. Осадки белков высушивают в вакуум-эксикаторе. [23]
 |
Гистограммы распределения и.с.у. нефтей. [24] |
Данные об и.с.у. давно широко используются в практике геохимических работ. Однако причины разного и.с.у. нефтей и их фракций до сих пор не ясны. Для объяснения наблюдаемых различий привлекают влияние типа исходного ОВ, процессов миграции, биодеградации, катагенеза. Предполагает, что ОВ скелетных форм планктона в значительной мере защищено от микробиологической атаки в зоне диагенеза, и поэтому большая доля легко разрушающихся и обогащенных тяжелым изотопом углерода белково-углеводных веществ фоссилизируется и принимает участие в последующих процессах нефтеобразования. Таким образом, находит объяснение существование в природе нефтей со сравнительно тяжелым и.с.у., которые незначительно отличаются от исходного морского ОВ. В отличие от скелетных бесскелетные формы планктона в ходе диаганеза теряют основную массу лабильных белков и углеводов, обогащенных тяжелым изотопом углерода, вследствие чего в образовании керогена ( а затем и нефтей) принимают участие другие, обогащенные легким изотопом углерода, компоненты исходного ОВ. Эта гипотеза существенно конкретизирует схему фракционирования и.с.у. на пути от исходного ОВ к нефти, однако она не позволяет объяснить большие различия и.с.у. нефтей, где вклад ОВ скелетных форм в общий баланс органического углерода мал. [25]
У эука-риот генетическая информация, содержащаяся в ядре, распределена между хромосомами. Каждая хромосома - это нитевидная структура, содержащая ДНК, основные белки особого типа, называемые гистонами и группу негистоновых белков, которые, вероятно, играют какую-то роль в регулировании функции генов. В неделящемся, или интерфазном, ядре каждая хромосома сильно выгнута и имеет толщину всего 20 - 30 нм; поэтому ее нельзя увидеть с помощью светового микроскопа. Интерфазное ядро содержит ядрышко - органеллу, богатую РНК и связанную со специфическим участком хромосомы - ядрышковым организатором. Ядрышковый организатор содержит множество копий генов, определяющих структуру рибосомальных РНК; ядрышко служит местом синтеза высокомолекулярного РНК-предшественника, из которого затем путем расщепления образуются основные типы молекул РНК, входящих в состав цитоплазматических рибосом. Эти РНК, а также матричные РНК, синтезируемые в других участках хромосом, выходят через ядерные поры в цитоплазму, где происходит сборка рибосом и синтезируется основная масса клеточного белка. [26]
Страницы: 1 2