Пурпурные бактерия
Cтраница 4
Как уже указывалось, убыль кислородного поглощения у пурпурных бактерий на свету Накамура считает доказательством фотохимического образования кислорода. Однако есть более вероятное объяснение: если скорости фотосинтеза и дыхания лимитированы наличием водорода, доставляемого одной и той же энзиматической системой, то всякое возрастание фотосинтеза будет подавлять дыхание. Тот факт, что эти организмы ни при каких обстоятельствах не переходят от поглощения кислорода к его выделению, хорошо согласуется с таким объяснением, тогда как он трудно объясним, если фотосинтез и дыхание независимы, как у высших растений. Каждая жирная кислота разлагается пурпурными бактериями с особой, характерной для нее, скоростью, одинаковой и для дыхания и для фотосинтеза; в случае смеси кислот их суммарная скорость разложения аддитивна. Это доказывает, что для каждой кислоты существует специфический энзнм. [46]
Эти исследователи нашли, что фоторедукция двуокиси углерода пурпурными бактериями ( Chromatium D) так же чувствительна к цианиду, как и фотосинтез Chlorella. [47]
Сероводород не является единственным восстановителем, который могут использовать пурпурные бактерии; в отличие от высших растений, их метаболизм чрезвычайно адаптивен. Некоторые виды предпочитают специфические восстановители, тогда как другие могут использовать в одинаковой мере целый ряд водородных доноров. Поэтому возможна лишь приблизительная классификация пигментированных бактерий с точки зрения их нормальной фотосинтетической функции. [48]
Несмотря на аналогию в обмене веществ адаптированных водорослей и пурпурных бактерий, он играет различную роль в жизни этих организмов: Гаффрон [17] обнаружил, что после нескольких дней фоторедукции у водорослей не проявлялось увеличения в содержании хлорофилла, сравнимого с увеличением, происходящим за такой же период фотосинтеза. [49]
Френч пришел к заключению, что красные каротиноидные пигменты пурпурных бактерий фотосинтетически неактивны. Следует отметить, что со спектроскопической точки зрения пурпурные бактерии особенно удобны для изучения роли каротиноидов благодаря тому, что максимумы поглощения бактериальных каротиноидов не закрываются полосами поглощения хлорофилла, как у зеленых растений и даже у фукоксантоловых водорослей. [50]
Не менее заманчивые перспективы открывает и обнаруженное сходство фотосинтезирующих пурпурных бактерий с некоторыми бесцветными бактериями, которые способны восстанавливать углекислоту с помощью тех же или близких восстановителей, но без участия света. Вместо энергии света они используют химическую энергию, образующуюся при ферментативном окислении этих восстановителей кислородом воздуха. Такой процесс называется бактериальным хемосинтезом. Возможно, что он также представляет собой реликт более древних форм жизни. В 1939 г. Гаффрон обнаружил, что некоторые одноклеточные зеленые водоросли, будучи лишены кислорода, перестают осуществлять обычный фотосинтез, но могут восстанавливать углекислоту на свету, если вместо обычного восстановителя - воды - обеспечить их водородом. Создается впечатление, как будто отсутствие кислорода вынуждает эти водоросли подражать пурпурным бактериям, которые также могут использовать в качестве восстановителя водород. [51]
НО) было указано, что при использовании фотосинтезирующими пурпурными бактериями подводимых извне органических соединений количество совместно ассимилируемой двуокиси углерода может колебаться в широких пределах ( двуокись углерода может даже выделяться) в зависимости от того, используется ли органическое вещество главным образом или даже исключительно в качестве донора водорода ( как в опытах Фостера со вторичными спиртами) или оно служит также источником углерода. Вассинк, Катц и Доррештейн полагают, что те же соображения применимы и к фотохимическому использованию внутриклеточных органических веществ; в этом случае потребление внешней двуокиси углерода также может быть более или менее полно подавлено использованием углерода ( в виде свежеобразованной двуокиси углерода или промежуточных продуктов окисления), образуемого при дегидрировании органического восстановителя. [52]
 |
Пути окисления соединений серы фототроф-нымн бактериями АФС - адепилилсульфат. [53] |
Эти данные весьма важны, поскольку до сих пор деление пурпурных бактерий на серные и несерные основывалось на их способности окислять сероводород. Сейчас такой принцип оказывается неприемлемым. [54]
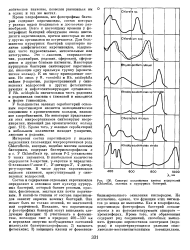 |
Спектры поглощения клеток водорослей ( Chlorella, зеленых и пурпурных бактерий. [55] |
Состав и содержание отдельных каротиноидов определяют в основном цвет культур пурпурных бактерий, который бывает розовым, красным, фиолетовым, желтым или почти коричневым. В какой-то степени от состава каротиноидов зависит окраска зеленых бактерий. [56]
Из различий в спектрах поглощения зеленых водорослей, цианобак-терий, пурпурных бактерий и зеленых бактерий можно заключить, что разные группы фототрофных организмов используют для фотосинтеза свет разных участков спектра. Это связано с условиями освещения в естественных местах обитания разных фототрофных организмов. [57]
Страницы: 1 2 3 4