Мембрана - тилакоид
Cтраница 3
Различия в структуре молекул хлорофилла приводят к тому, что каждый из них обладает уникальным спектром поглощения. Внутри группы молекул хлорофилла а есть различия в светопог-лощении молекул, которые определяются конкретным микроокружением в липопротеиновом слое мембраны тилакоидов. Существуют достоверные данные о наличии двух специализированных видов молекул хлорофилла а, максимумы поглощения которых соответствуют 680 и 700 нм. [31]
Мембраны, в которых локализованы ферменты дыхательной цепи и окислительного фосфорилирования, называют сопрягающими мембранами. Примерами таких мембран являются внутренняя мембрана митохондрий, клеточная мембрана аэробных бактерий с дыхательным типом энергетики, хроматофоры фотосинтезирующих бактерий и мембраны тилакоидов хлоропластов зеленых растений. Отличительным признаком сопрягающих мембран является их способность образовывать АТФ за счет энергии внешних ресурсов. [32]
На поверхности мембран тилакоидов упорядоченными рядами располагаются выступающие структуры - партикулы, неодинаковые по величине и плотности. С наружной стороны мембран тилакоидов, в зоне, соприкасающейся со стромой, размещены партикулы размером около 11 нм, с которыми структурно связана ФС I. В процессе выделения они оказываются в одной фракции со стромой. [33]
В описанной модели световой стадии фотосинтеза каждый электрон пересекает мембрану тилакоида дважды. Такое перемещение сопровождается исчезновением из стромы трех протонов и появлением внутри тилакоида четырех протонов. Перемещение протонов переводит мембрану тилакоида в знергизованное состояние. [34]
Фотосинтезирующие реакционные центры ( ловушки экситонов) входят в состав фотосинтезирующих систем ( ФС), в которых осуществляются световые реакции фотосинтеза. Фотосинтезирующие системы наряду с реакционными центрами, воспринимающими энергию света, содержат ряд других молекул - ферменты, белки, липиды, липопротеиды, которые участвуют в организации фото-синтезирующей системы и в выполнении ею световой части биохимических реакций. Фотосинтезирующие системы сравнительно жестко вмонтированы в мембраны тилакоидов. [35]
Хемиосмотическая гипотеза дает объяснение ряду экспериментальных фактов. При освещении хлоропластов наблюдается иовышение рН раствора ( внутри тилакоида наблюдается подкис-ление), обратимое в темноте и чувствительное к ингибиторам электронного транспорта. Создание в темноте искусственного градиента протонов на мембранах тилакоидов сопровождается синтезом молекул АТФ. [36]
Мембраны тилакоидов являются основными сопрягающими мембранами хлоропластов. По-видимому, молекулы хлорофилла располагаются со стороны внутренней поверхности мембраны тилакоида. В живых клетках полярность мембран тилакоидов обратна полярности сопрягающих мембран митохондрий. У тилакоидов внутренняя поверхность мембраны имеет положительный электрический потенциал, а внешняя - отрицательный. [37]
В свою очередь молекулы цитохрома /, получив электроны от молекул PQ Н2, передают их дальше, к белковым молекулам, содержащим атомы меди и называемым пластоцианидами. Молекулы пластоцианида передают электроны реакционным центрам Рт первой фотосистемы. Электроны же, оторвавшиеся от этих центров при поглощении света, перемещаются к внешней поверхности мембраны тилакоида и захватываются железо-серными белками, от которых они переходят к молекулам ферредоксина, а от них - к молекулам ФАД. [38]
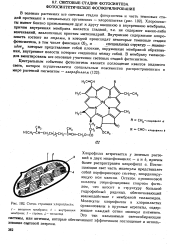 |
Схема строения хлоропласта. [39] |
Хлорофилл встречается у зеленых растений в двух модификациях - а и Ь, причем более распространен хлорофилл а. Поглощающая свет часть молекулы представляет собой порфириновую систему, координирующую ион магния. Остаток пропионовой кислоты этернфицирован спиртом фито-лом, что вносит в структуру большой гидрофобный радикал, обеспечивающий взаимодействие с мембраной тилакоидов. Молекулы хлорофилла в хлоропластах организованы в крупные агрегаты, содержащие каждый сотни молекул пигмента. Это так называемые светособирающие системы, или антенны, которые обеспечивают эффективное поглощение и использование световой энергии. [40]
Мембраны тилакоидов являются основными сопрягающими мембранами хлоропластов. По-видимому, молекулы хлорофилла располагаются со стороны внутренней поверхности мембраны тилакоида. В живых клетках полярность мембран тилакоидов обратна полярности сопрягающих мембран митохондрий. У тилакоидов внутренняя поверхность мембраны имеет положительный электрический потенциал, а внешняя - отрицательный. [41]
Далее в цепи переноса электронов от фотосистемы 11 к фотосистеме 1 принимают участие два цито. Следующим компонентом цепи является медьсодержащий белок пластоцианин, от которого электрон поступает в реакционный центр фотосистемы I. Комплекс, образованный нластохиноиом, цитохромами 6559 и С552 и пластоцнанином, по-видимому, является близким аналогом комплекса III в цепи переноса электронов в митохондриях. При прохождении через него пары электронов создается градиент концентрации протонов через мембрану тилакоидов, направленный внутрь мембраны, - рН внутри тила-коида может достигать значений 4 и ниже. [42]
Согласно хемиосмотической гипотезе, синтез молекул АТФ в тилакоиде осуществляется комплексом ферментов CF1 - F0, эквивалентным комплексу ферментов F1 - F0, входящему в состав сопрягающих мембран митохондрий. Ферменты CF1 находятся на внешней стороне мембраны тилакоида, образуя шишковидные выступы. Такое расположение обусловлено тем, что направление протонного градиента в тилакоидах противоположно направлению градиента протонов в мембранах бактерий и сопрягающих мембранах митохондрий. Интересно, что в липосомах, образованных в суспензиях из фрагментов мембран митохондрий, ферменты GFj выступают на внешней поверхности мембраны пузырьков так же, как и в естественных мембранах тилакоидов. [43]
Механизм фотосинтетического фосфорилирования сходен с синтезом АТФ в процессе окислительного фосфорилирования в митохондриях. Система переносчиков электронов интегрирована в мембрану тилакоида таким образом, что перенос пары электронов создает поток протонов с наружной поверхности тилакоида внутрь, рН на внутренней поверхности тилакоида может достигать 4 и ниже. Структура этого фермента аналогична митохондриальной АТФ-синтетазе ( гл. CFr Символ С означает, что этот ферментный комплекс локализован в хлоропластах ( chloroplasf) и, подобно митохондриальной Н - зависимой - АТФ-синтетазе, включает гидрофобный, интегрированный в мембрану тилакоида компонент ( CF0) и гидрофильный комплекс ( CFj), катализирующий синтез АТФ. [44]
Существует два основных процесса энергообеспечения таких систем: фотосинтез и дыхание. У высших организмов процессы энергообеспечения локализованы на мембранах особых компонентов клетки ( внутриклеточных органелл) - митохондрий, которые служат своеобразными силовыми станциями, поставляющими энергию, необходимую для функционирования клеток. Митохондрия окружена двумя мембранами - внешней и внутренней. Ферментные комплексы, ответственные за энергообеспечение клетки, расположены на внутренней мембране митохондрий. Аналогичную роль играет клеточная мембрана аэробных бактерий. У зеленых растений трансформация энергии происходит на мембранах тилакоидов хлоропластов, а у фотосинтезирующих бактерий - на мембранах хроматофоров. Увеличение ионной проводимости мембран приводит к рассеиванию энергии в виде теплоты, а разрушение мембран - к полной потере способности к аккумуляции энергии. [45]
Страницы: 1 2 3 4