Тилакоидная мембрана
Cтраница 2
Эти незаменимые жирные кислоты очень важны для формирования связи между хлорофиллом и тилакоидной мембраной, а также для сохранения структуры гран. Определенные гербициды трансформируют линоленовую кислоту в перекись ( а-линолен ООН), которая вызывает окисление хлорофилла. [16]
В то же время можно рассчитать, пользуясь электронными микрофотографиями, как велика общая поверхность всех тилакоидных мембран. Оказывается, что обе полученные величины поразительно точно совпадают. [17]
Выделение энергии при превращениях АДФ - АТФ и фотовозбуждение при фотофосфорилировании приводят к возникновению разности концентраций водородных ионов внутри и вне тилакоидной мембраны. При освещении обычным светом изменение рН приблизительно равно 3, что энергетически недостаточно для осуществления требуемых процессов. [18]
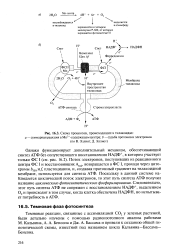 |
Схема процессов, происходящих в тилакоидах. а - суммарная реакция в Мп2 - содержащем центре. б - судьба протонов и электронов. [19] |
Поток электронов, поступивших из реакционного центра ФС I и восстановивших А, возвращается в ФС I, проходя через цито-хромы Z559 и /, пластоцианин, и, создавая протонный градиент на тилакоидной мембране, используется для синтеза АТФ. Поскольку в данной системе наблюдается циклический поток электронов, то этот путь синтеза АТФ получил название циклического фотосинтетического фосфорилирования. Следовательно, этот путь синтеза АТФ не сопряжен с восстановлением НАДФ, выделением 02 и происходит в том случае, когда клетка обеспечена НАДФН, но испытывает потребность в АТФ. [20]
Основные фотобиологические процессы - фотосинтез и фоторецепция - разыгрываются в мембранах. Тилакоидные мембраны хлоропластов, мембраны фоторецепторных клеток имеют специфическое строение и функции, еще далеко не познанные. [21]
Сине-зеленые водоросли ( Cyanophyta или СуапоЬас-teria) представляют собой единственную большую группу прокариот, которые способны к фотосинтезу с выделением кислорода, сходному с фотосинтезом у высших растений. Однако тилакоидные мембраны у них находятся не в хлоропластах, а распределены по всей цитоплазме клетки, преимущественно на ее периферии. [22]
Фотосистема II, максимально активируемая светом с длинами волн короче 680 нм, содержит относительно больше хлорофилла Ь, а иногда также хлорофилл с. В тилакоидных мембранах одного хлоропласта шпината имеется много сотен фотосистем того и другого типа. Позднее мы увидим, что эти две фотосистемы выполняют разные функции. [23]
Окончательное состояние тилакоидных мембран зависит от условий окружающей среды, главным образом от освещения. Хлоропласты, развивавшиеся при высоких интенсивностях освещения, имеют относительно небольшие, но высокоэффективные ФС I и ФСП, тогда как ССК У них редуцированы. При более низких интенсивностях освещения, при которых поглощение света должно быть по возможлости максимально эффективным, большое значение имеет синтез ССК, связанный с организацией тилакоидов в граны. Имеются сообщения, что первичными регуляторами развития хлоропластов служат красный свет и фитохромнаЯ система. Однако, согласно другим сообщениям, важную роль в данном случае играет синий свет и пока еще неизвестный фоторецептор. [24]
Помимо хлорофилла, в процессах фотосинтеза участвуют пигменты группы каротиноидов, в состав которых входят только водород и углерод, и ксантофиллы, имеющие в составе молекул еще и кислород. Пигменты встречаются в тилакоидных мембранах всех фотоавтотрофных организмов. Каротиноиды играют роль антенных пигментов, чувствительных к солнечному свету в диапазоне волн, недоступном для хлорофилла. Они передают энергию солнечного света в центры реакций и, кроме того, как светофильтры экранируют хлорофилл в листьях, предохраняя его от фотодеструкции, фотоокисления. Этот защитный эффект связывают с наличием конъюгированных двойных связей ( их может быть 9 или более), способных гасить возбужденное состояние молекул хлорофилла. Каротин - протеиновый комплекс Csso, расположенный в акцепторной части ФС II, также может участвовать в окислительно-восстановительных процессах. [25]
 |
Цикл Кальвина - превращение СО2 в глюкозу в процессе фотосинтеза. [26] |
Фермент - рибулозодифосфат-карбоксилаза имеет очень сложное строение и огромную молекулярную массу - 550 000 Да. Фермент локализуется на наружной поверхности тилакоидных мембран, он широко распространен в природе - это главный фермент, ответственный за образование биомассы растений из СО2 на нашей Земле. [27]
Фотобатареи жидкостного типа представляются перспективными в качестве аккумуляторов энергии. В таких батареях разделение зарядов осуществляется ( аналогично тилакоидным мембранам фотосинтетических систем) путем катодного восстановления и анодного окисления. Двухслойные молекулярные мембраны ( типа показанных на рис. 4.14) обладают малой прочностью, но их характеристики можно улучшить, используя в качестве подложек микропористые пленки. Очень важным является создание полимерных пленок, способных осуществлять разделение электрических зарядов в фотосинтетических системах. Такие пленки позволили бы регулировать обратные реакции в процессах, описанных в разд. [28]

В то же время поверхность, на которой расположены доноры электронов, недо - ступна для антител. Эту схему пространственной ориентации направленного фотосинтетического транспорта электронов внутри тилакоидной мембраны следует отличать от схемы на рис. 12 14, где компоненты расположены по вертикали в соответствии с их окислительно-восстановительными потенциалами. [30]
Страницы: 1 2 3 4