Электровозбудимая мембрана
Cтраница 1
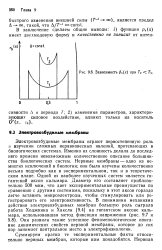 |
Зависимость р ( х при Т2 Т. [1] |
Электровозбудимые мембраны играют первостепенную роль а изучении сложных неравновесных явлений, протекающих в биологических системах. Именно их сложность делала до последнего времени невозможным количественное описание большинства биологических систем. Нервные мембраны - одно из немногих исключений в биологии; они были изучены количественно весьма подробно как в экспериментальном, так и в теоретическом плане. Одной из наиболее изученных систем является гигантский аксон кальмара. Диаметр его составляет приблизительно 500 мкм, что дает экспериментальные преимущества по сравнению с другими системами, поскольку в этот аксон гораздо легче вставить микроэлектроды, чтобы стимулировать или регистрировать его электроактивность. Они развили также успешное феноменологическое описание динамических свойств нервной мембраны, которое до сих пор занимает центральное место в электрофизиологии. [2]
Другой формой пластичности электровозбудимой мембраны является смещение порога генерации ПД. Порог генерации характеризуется тем значением уровня мембранного потенциала, который должен быть достигнут, чтобы начался лавинообразный процесс генерации ПД. Повышение порога генерации ПД означает смещение этого уровня в направлении нуля. Снижение порога означает сдвиг этого значения в сторону большей поляризации. При повышении порога приходящие ВПСП перестают его достигать, при снижении - даже ВПСП низкой амплитуды генерируют ПД. [3]
Нарушение связи между микроучастками электровозбудимой мембраны приводит к тому, что цепная реакция либо вообще не возникает, либо развивается в ограниченном участке электровозбудимой мембраны. [4]

Между деполяризованной ацетилхолином постсинаптической мембраной и соседними с ней участками электровозбудимой мембраны скелетного мышечного волокна возникают местные токи, вызывающие генерацию потенциала действия, распространяющегося по всему мышечному волокну. Условием возникновения этого потенциала действия является критическая деполяризация электровозбудимой мембраны, происходящая при достижении ПКП пороговой величины. [6]
 |
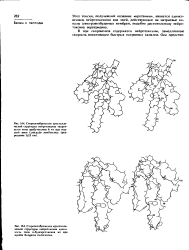
Структура токсина II из яда средиземноморской анемоны Anemonia sulcata. [7] |
Подобно нейротоксинам яда скорпиона, они действуют на быстрые натриевые кана1ы электровозбудимых мембран, влияя на процесс инактивации. Однако по структурным признакам токсины аиемон достаточно сильно отличаются от скорпионовых токсинов. На рисунке 168 представлена структура типичного представителя этой группы гомологичных полипептидов - токсина 11, выделенного из средиземноморской анемоны Anemonia sulcata. Токсин II состоит из 47 аминокислотных остатков с тремя внутримолекулярными днсульфнднымн связями и не имеет видимых структурных аналогий ни с токсинами яда змей, ни с токсинами скорпионов. Для этого нейротоксина следует отметить присутствие большого числа гидрофобных аминокислотных остатков. [8]
Этот токсин, получивший название кротамин, является единственным нейротоксииом яда змей, действующим на натриевые каналы электровозбудимых мембран, подобно растительному нейро-токсину вератридину. [10]
Можно предположить, что волна деполяризации, образующая колебания пейсмекерного потенциала, представляет собой медленно развивающийся процесс электровозбудимой мембраны, аналогичный по механизму ПД. Известно, что генерация ПД связана с открыванием натриевых пор. Вход натрия в нейрон по электрохимическому градиенту блокируется тетродотоксином, переводящим натриевые каналы в латентное состояние. В результате ПД полностью исчезает. [11]
Токсин снижает уровень поляризации физических мотонейронов за счет повышения проницаемости их поверхностных мембран, вероятно инактивируются натриевые каналы электровозбудимой мембраны. [12]
Изменение порога генерации ПД под влиянием процедуры обусловливания в изолированном командном нейроне заставляет высказать предположение, что в этом процессе замешана электровозбудимая мембрана. [13]
Можно предположить, что в росте ответа на апликацию АХ при подкреплении его током, вызывающим спайки, также участвуют локусы электровозбудимой мембраны, увеличивая деполяризационный эффект действия АХ. [14]
Нарушение связи между микроучастками электровозбудимой мембраны приводит к тому, что цепная реакция либо вообще не возникает, либо развивается в ограниченном участке электровозбудимой мембраны. [15]
Страницы: 1 2