Метаболизм - мозг
Cтраница 1
Метаболизм мозга примечателен в ряде отношений. Во-первых, в качестве клеточного топлива мозг взрослых млекопитающих обьино использует только глюкозу. Во-вторых, мозгу свойствен очень интенсивный дыхательный обмен; так, у взрослого человека мозгом используется почти 20 % кислорода, потребляемого организмом в состоянии покоя. Кроме того, скорость использования кислорода мозгом отличается большим постоянством и не претерпевает существенных изменений при переходе, например, от активной умственной работы ко сну. Из-за низкого содержания гликогена мозг нуждается в постоянной доставке глюкозы кровью. [1]
Поскольку метаболизм работающего мозга требует подпитки, он потребляет больше крови. За этими процессами следят датчики радиоактивности, данные от которых преобразуются компьютером в географическую карту коры мозга, по которой можно определить горячие точки его отделов, где сосредоточено больше крови. [2]
Говоря о нарушениях белкового метаболизма мозга, вызванных пренатальной алкоголизацией, следует иметь в виду, что эти нарушения касаются всех видов белков, в том числе ферментных. Понятно, что это не может не сказаться на тех метаболических звеньях, в которых участвуют соответствующие ферменты. [3]
Итак, воздействие алкоголя в пренаталъном периоде приводит к нарушениям белкового метаболизма мозга в постнаталъном периоде. Эти нарушения могут быть явными, легко выявляемыми, но могут носить и скрытый характер, обнаруживаясь лишь при использовании специальных приемов. [4]
Наиболее существенные изменения у подопытных животных были выявлены при исследовании белкового метаболизма мозга. Соответствующие данные представлены на рис. 9, где показаны особенности включения 358-метио-нина в белки церебральных структур 2-го поколения подопытных животных разного возраста. [5]
Проникающий в мозг зародыша, эмбриона и плода этанол, изменения метаболизма развивающегося мозга и другие факторы, связанные с алкоголизацией родителей, в том числе изменения половых клеток и вызванные хронической алкоголизацией изменения генетического материала, в совокупности приводят к нарушениям развития ЦНС, ее врожденным дефектам, составляющим главный компонент АСП и алкогольных эмбриопатий. В основе этих дефектов, как будет видно из дальнейшего, лежат задержки развития нервных клеток и мозга в целом, нарушения связей между нервными клетками, глубокие нарушения метаболизма мозга и его микро - и макроструктуры. Существенно, что в происхождении задержек развития нервных клеток и мозга в целом у потомства алкоголизированных животных важную роль играют нарушения метаболизма белка - угнетение его синтеза. [6]
Если ограничиться этими показателями, то легко прийти к заключению о полноценности белкового метаболизма мозга у подопытных животных. [7]
 |
Схема превращения глюкозамина. [8] |
Результаты исследования убеждают нас в лабильности глюко-замина и возможности его активного участия в метаболизме мозга. [9]
Попятно, что за счет повреждения мембран митохондрий может нарушаться главная функция этих органоидов, что, естественно, приводит к усугублению гипоксии и снижению энергетического обеспечения всех, в том числе биосинтетических, процессов в клетке. В результате глубоко нарушается метаболизм развивающегося мозга, что и приводит к нарушениям функций высших отделов ЦНС в постпаталь-ном периоде. [10]
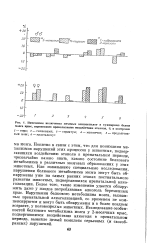 |
Изменения включения меченых аминокислот в суммарные белки мозга крыс, перенесших пренатальные воздействия этанола, % к контролю. [11] |
Понятно в связи с этим, что для понимания механизмов нарушений этих процессов у животных, подвергавшихся воздействию этанола в пренатальном периоде, чрезвычайно важно знать, каково состояние белкового метаболизма в различных мозговых образованиях у этих животных. Как показывают специальные исследования, нарушения белкового метаболизма мозга могут быть обнаружены уже на самых ранних этапах постнатального развития животных, подвергавшихся пренаталъной алкоголизации. Более того, такие изменения удается обнаружить даже у плодов потреблявших алкоголь беременных крыс. Нарушения белкового метаболизма мозга, вызванные пренаталъной алкоголизацией, со временем не компенсируются и могут быть обнаружены и в более позднем возрасте, у половозрелых животных. [12]
Несмотря на наличие обширной литературы о действии алкоголя на потомство, ряд аспектов этой проблемы освещен недостаточно. Это касается, в частности, функции, структуры и метаболизма мозга животных, подвергавшихся действию алкоголя во внутриутробном периоде, Между тем без знания механизмов возникновения таких нарушений невозможна разработка конкретных методов терапии и профилактики врожденных поражений мозга алкогольного генеза. [13]
Разная степень поступления радиофосфора в различные мозговые структуры отражает неодинаковую степень изменения проницаемости сосудов этих структур. Как известно, проницаемость ГЭБ для веществ, участвующих в метаболизме мозга, определяется метаболическими запросами соответствующих мозговых образований и зависит от уровня их функциональной активности. [14]
Многими исследователями выдвигается представление о том, что единым патофизиологическим механизмом, через который реализуется болезнетворное влияние различных патогенных факторов на внутриутробно развивающийся организм, является гипоксия. Если это положение справедливо, то представляется весьма целесообразным сопоставление особенностей структуры, функции и метаболизма мозга животных, подвергавшихся в пренаталь-ном периоде воздействию чистой гипоксии и алкоголя. [15]
Страницы: 1 2