Аэробный метаболизм
Cтраница 2
Скорее, следует сказать, что новый механизм наложился на старый, дополнив и улучшив его. Аэробный метаболизм вступает в дело там, где перестает действовать анаэробный метаболизм. [16]
В подготовленное таким образом сусло добавляют дрожжевые клетки. В аэробном сусле дрожжи растут и размножаются очень быстро, извлекая необходимую им энергию из некоторых присутствующих в сусле Сахаров. Аэробный метаболизм дрожжей обусловливает очень быстрый рост клеток, регулируется же этот метаболизм добавлением нужного количества кислорода. После исчерпания всего растворенного кислорода в чане с суслом дрожжевые клетки как факультативные анаэробы ( разд. Процесс брожения регулируется концентрацией образовавшегося этанола, а также величиной рН и количеством несброженного сахара. В определенный момент брожение останавливают, удаляют дрожжи, и молодое, или зеленое, пиво поступает на дображивание. Светлое пиво, которое стало теперь очень популярным, содержит меньше сахара и алкоголя, чем обычное, однако по своему аромату оно не отличается от обычных сортов. [17]
Исчерпание молекулярного кислорода in situ приводит к замедлению тепловыделения, поступление кислорода за счет конвекции также соответственно снижается. Одновременно накопление диоксида углерода в течение стадии компостирования создает микроаэрофильные условия, которые приводят к увеличению числа сначала факультативных, а затем и облигатных анаэробов. В отличие от аэробного метаболизма, при котором минерализация отходов часто достигается с помощью одного вида бактерий, анаэробная биодеградация требует совместного метаболизма микроорганизмов разных видов, входящих в состав смешанной популяции. Эта популяция взаимодействующих друг с другом микроорганизмов способна использовать различные неорганические акцепторы электрона, часто в последовательности, соответствующей выделению энергии при этой реакции. Так как большинство бактерий нуждается в определенных акцепторах электронов, то эта последовательность приводит к существенным изменениям в составе микробной популяции. Виды, способные использовать более окисленные акцепторы, получают термодинамические и, следовательно, кинетические преимущества. [18]
Вообще говоря, у мелких животных кислород доставляется к мышцам циркуляторными системами достаточно быстро, так что необходимости в анаэробном использовании мышечного гликогена у них нет. Птицы, например, при своих перелетах часто покрывают огромные расстояния с очень большой скоростью без какой бы то ни было кислородной задолженности. Красным мышцам многих бегающих животных среднего размера также свойствен по преимуществу аэробный метаболизм. Однако у крупных животных при напряженной и длительной работе циркуляторная система оказывается уже не в состоянии поддерживать полностью аэробный метаболизм в мышцах. Эти животные движутся обычно медленно, и только крайние обстоятельства вынуждают их к усиленной мышечной активности, поскольку за каждой такой вспышкой активности должен следовать долгий период восстановления, необходимый для погашения кислородной задолженности. [19]
 |
Влияние щелочности среды на рост микроорганизмов. а Pseudomonas 7 - 2Л - В. 6 - Pseudomonas aeruginosa. [20] |
Это окисление с выделением энергии происходит путем перехода протонов или электронов через ряд стадий, регулируемых ферментами, до появления конечного акцептора электронов. В аэробных процессах конечным акцептором электрона или иона водорода является кислород. В анаэробных процессах таким акцептором является окисленный материал типа нитрата или сульфата. Опыт показал, что аэробный метаболизм эффективнее анаэробного, так как для роста в аэробных процессах требуется меньше материала подложки, чем в анаэробных при одинаковом количественном росте бактерий. Причиной такого явления, известного как эффект Пастера, является большее выделение энергии в процессе аэробного метаболизма. [21]
Все эти дрожжи способны продуцировать значительные количества липидов. Однако в отличие от С - terficolus лини-дообразование их существенно зависит от словий к льтиииповяния и R перную очередь от соотношения соединений углерода и азота в среде. Общим для перечисленных микроорганизмов является строго аэробный метаболизм и неспособность к росту в результате брожений. [22]
Вообще говоря, у мелких животных кислород доставляется к мышцам циркуляторными системами достаточно быстро, так что необходимости в анаэробном использовании мышечного гликогена у них нет. Птицы, например, при своих перелетах часто покрывают огромные расстояния с очень большой скоростью без какой бы то ни было кислородной задолженности. Красным мышцам многих бегающих животных среднего размера также свойствен по преимуществу аэробный метаболизм. Однако у крупных животных при напряженной и длительной работе циркуляторная система оказывается уже не в состоянии поддерживать полностью аэробный метаболизм в мышцах. Эти животные движутся обычно медленно, и только крайние обстоятельства вынуждают их к усиленной мышечной активности, поскольку за каждой такой вспышкой активности должен следовать долгий период восстановления, необходимый для погашения кислородной задолженности. [23]
Тот факт, что при инкубации ткани печени с тироксином увеличивается интенсивность дыхания и теплообразования и не изменяется концентрация АТР, согласуется с утверждением, что тироксин является разобщителем окислительного фосфорилиро-вания. Разобщающие агенты понижают отношение Р / О в тканях; это заставляет ткани увеличивать интенсивность дыхания, чтобы удовлетворить потребность в АТР. Наблюдаемое выделение тепла могло быть обусловлено также повышением скорости утилизации АТР тканью, стимулированной тироксином. В такой ткани возросшая потребность в АТР удовлетворяется за счет повышения уровня окислительного фосфо-рилирования ( дыхания), что сопровождается выделением тепла. Несмотря на многочисленные исследования, детали регуляции тиреоидными гормонами скорости аэробного метаболизма остаются загадкой. [24]
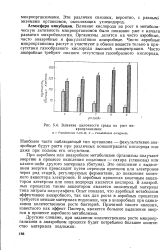 |
Влияние щелочности среды на рост микроорганизмов. а Pseudomonas 7 - 2Л - В. 6 - Pseudomonas aeruginosa. [25] |
Это окисление с выделением энергии происходит путем перехода протонов или электронов через ряд стадий, регулируемых ферментами, до появления конечного акцептора электронов. В аэробных процессах конечным акцептором электрона или иона водорода является кислород. В анаэробных процессах таким акцептором является окисленный материал типа нитрата или сульфата. Опыт показал, что аэробный метаболизм эффективнее анаэробного, так как для роста в аэробных процессах требуется меньше материала подложки, чем в анаэробных при одинаковом количественном росте бактерий. Причиной такого явления, известного как эффект Пастера, является большее выделение энергии в процессе аэробного метаболизма. [26]
В отношении простейших экологическая ситуация подвергается значительным изменениям. Среда, окружающая их, становится богатой углеродом, что благоприятствует росту простейших и закономерно ведет к утилизации кислорода. Интервал времени между ростовыми циклами определяется временным снижением потребления кислорода. Потребление кислорода в ходе метаболизма простейших представляет третью стадию, поскольку вторая стадия потребления кислорода представляет относительно низкую величину и если ее не учитывать, то потребление кислорода простейшими можно именовать второй углеродной стадией потребления кислорода, В реке простейшие в свою очередь становятся пищей для высших организмов. Этот аэробный метаболизм также оказывает влияние на потребление кислорода. [27]
Одной из основных проблем современной биохимии является выяснение механизма превращения энергии, выделяющейся в результате взаимодействия связей С - Н с кислородом с образованием двуокиси углерода и воды в энергию фосфоангидридной связи АТФ - единой платежной единицы в процессах переноса химической энергии, используемой для большого числа синтетических и метаболических функций. Если энергетическое сопряжение имеет химический механизм ( хотя это еще не очевидно [185]), то оно может происходить либо непосредственно через окисление некоторых легко образующихся низкоэнергетических фосфатных производных до высокоэнергетических форм, которые могут затем переносить фосфат на АДФ, давая АТФ, либо через окисление некоторых других низкоэнергетических молекул до высокоэнергетических форм, которые могут дать макроэргический фосфат через серию реакций переноса. В последнее время стало известно несколько примеров такого актива-ционного процесса, в котором происходит образование высокоэнергетического тиолового эфира при окислении альдегида. Тиоловый эфир может реагировать дальше, давая ацилфосфат и при известных обстоятельствах АТФ. Этот тип активации является ответственным за образование макро-эргических фосфатных связей на субстратном уровне фосфорилирования, в котором метаболит, подвергающийся окислению, превращается в активированный продукт. В настоящее время, однако, еще нет уверенности, что аналогичный процесс происходит при многоступенчатом переносе электронов между субстратом и кислородом, который является ответственным за освобождение большей части энергии в аэробном метаболизме. Интерес к этой проблеме стимулировал поиски реакций, в которых фосфатная группа превращается в энергетически богатую форму посредством окислительного процесса, что может служить моделью реакций с природным коферментом. Хотя в настоящее время еще нет доказательств, что какой-либо процесс такого рода ответствен за окислительное фосфорилирование, эти исследования интересны с химической точки зрения и в качестве источника некоторых потенциально полезных синтетических методов. [28]
Страницы: 1 2