Энергетический метаболизм
Cтраница 3
ЦТК, активное флавиновое дыхание и окислительное фосфорилирование, сопряженное с электронтранспор-тной системой. Вклад каждого из этих путей в общий энергетический метаболизм зависит как от вида бактерий, так и от конкретных внешних условий. [31]
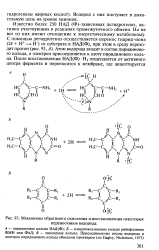 |
Механизмы обратимого окисления и восстановления некоторых переносчиков водорода. [32] |
Известно более 250 НАД ( Ф) - зависимых дегидрогеназ, активно участвующих в реакциях промежуточного обмена. Но не все из них имеют отношение к энергетическому метаболизму. Атом водорода входит в состав пиридинового кольца, а электрон присоединяется к азоту пиридинового кольца. [33]
Свойство предельной окисленности молекулы С02 используется в энергетическом метаболизме ряда анаэробных эубактерий, получающих энергию в процессе брожения, где С02 служит для удаления избытка восстановителя, т.е. как конечный акцептор электронов. Эта же особенность молекулы С02 находит применение и в энергетическом метаболизме некоторых анаэробных эубактерий ( ацетогены) и архебактерий ( метаногены), но у них электроны на С02 поступают через цепь связанных с мембраной переносчиков электронного транспорта. В наибольшей степени способность вовлекать С02 в метаболизм среди первично анаэробных хемогете-ротрофных эубактерий проявляется в группе клостридиев. [34]
Хлебопекарные дрожжи обладают и бродильной активностью, но чтобы достигнуть использования Сахаров только для образования биомассы, спиртовое брожение надо ограничить всеми доступными средствами. При высокой концентрации Сахаров имеет место катаболигная репрессия ферментов цикла Кребса и переключение энергетического метаболизма преимущественно на брожение. Чтобы избежать этого, сахар в среду подают непрерывно с постоянной или возрастающей скоростью притока. [35]
Организмы, у которых источниками ( донорами) электронов в энергетическом процессе являются неорганические вещества, предложено называть литотрофны-м и, а те, у которых донорами электронов служат органические соединения, - органотрофными. Тогда в зависимости от источника энергии и природы донора электронов возможны четыре основных типа энергетического метаболизма: хемолитотро-фия, хемоорганотрофия, фотолитотрофия и фо-тоорганотрофия. [36]
НАД в качестве кофермента; А - молекула органического вещества, служащая акцептором электронов. Молекулы НАДН2 используются в конструктивном метаболизме, обеспечивая восстановитель для биосинтетических процессов, а также в системе энергетического метаболизма, участвуя в решении акцепторной проблемы. Электронный перенос в этом случае не приводит к получению ютет-кой энергии, она вырабатывается только в реакциях субстраткюго фосфорилирования. [37]
То, как эта возможность реа-лизовывалась, доказывает наличие нескольких типов фотосинтеза, осуществляемого разными группами эубактерий, энергетический метаболизм которых полностью или частично основан на использовании энергии света. Фотосинтезирующие эубактерий представлены пурпурными и зелеными бактериями, гелио-бактериями, цианобактериями1 и прохлорофитами. [38]
Эти роды объединены по определителю бактерий Берги в трибу Slreptococcaceae, семейство Lactobacillaceae. Общие признаки перечисленных выше родов: грамположительные, образуют цепочки, неподвижные, кокки, микро-аэрофилы или факультативные анаэробы, ка-талазу не образуют, энергетический метаболизм протекает по типу брожения, нуждаются в сложных питательных веществах. [39]
Воздух как среда жизни обладает определенными особенностями, направляющими общие пути эволюции обитателей этой среды. Так, высокое содержание кислорода ( около 21 % та атмосферном воздухе, несколько меньше - в воздухе, заполняющем дыхательную систему животных) определяет возможность формирования высокого уровня энергетического метаболизма. Не случайно именно в этой среде возникли гомойотермные животные, отличающиеся высоким уровнем энергетики организма, большой степенью автономности от внешних воздействий и высокой биологической активностью в экосистемах. С другой стороны, атмосферный воздух отличается низкой и изменчивой влажностью. [40]
 |
Типы анаэробного дыхания у эубактерий. [41] |
Эубактерий, у которых источником энергии служат процессы окисления неорганических соединений, были обнаружены в конце XIX в. Разнообразие наблюдается только на периферических участках энергетического метаболизма, так как для окисления неорганических соединений, связанного с получением энергии, необходимы соответствующие ферментные системы. [42]
У многих эубактерий конечным акцептором электронов дыхательной цепи, наиболее часто заменяющим молекулярный кислород, служит нитрат. Он может восстанавливаться до нитрита, накапливающегося в среде, или молекулярного азота, удаляющегося в атмосферу. Процесс восстановления нитрата до нитрита в системе энергетического метаболизма, получивший название н и-тратного дыхания, широко распространен среди эубактерий и обнаружен у представителей более 70 родов. [43]
В течение многих лет покоящаяся спора рассматривалась как почти инертное тело, обладающее несколькими ферментами или вовсе лишенное их. Некоторых ферментных систем, имеющихся в вегетативной клетке, например цитохромов и ферментов цикла трикарбоновых кислот, ничтожно мало в споре. Это указывает на то, что в спорах аэробный энергетический метаболизм осуществляется иным путем, чем в клетках. Аналогичные ферменты спор и вегетативных клеток в спорах более термоустойчивы. [44]
Существуют разные точки зрения относительно роли мезосом в клетке. Согласно одной из них мезосомы не являются обязательной структурой, а служат только для усиления определенных клеточных функций, увеличивая общую рабочую поверхность мембран. Получены данные о том, что с мезосомами связано усиление энергетического метаболизма клеток. Мезосомы играют роль в репликации хромосомы и ее последующем расхождении по дочерним клеткам, участвуют в процессе инициации и формирования поперечной перегородки при клеточном делении. Для некоторых грамположительных бактерий обнаружено участие мезосом в секреторных процессах. [45]
Страницы: 1 2 3 4