Миграция - заряд
Cтраница 3
Алкилирование протекает по карбоний-ионному механизму. На четвертой стадии происходит перегруппировка этого иона вследствие миграции заряда вдоль углеродной цепи. Пятая, последняя стадия - взаимодействие этого иона с изопарафином по третичной углерод-водородной связи с образованием конечных продуктов реакции и новых карбоний-ионов, способных продолжить цепь. [31]
Наиболее достоверным, видимо, является механизм, предложенный Шмерлингом. На четвертой стадии происходит перегруппировка этого иона за счет миграции заряда вдоль углеродной цепи. Пятая, последняя стадия - взаимодействие этого иона с изопарафином по третичной углерод-водородной связи с образованием конечных продуктов реакции и новых карбоний-ионов, способных продолжить цепь. [32]
В последнем разделе собрано несколько статей, посвященных изучению водородных связей в биологических объектах. В частности, первая из этих статей представляет собой обзор исследований миграции зарядов в белках; как при протонном, так и при электронном механизме миграции водородные связи играют существенную роль. [33]
Оператор М связан с разностью электронных дипольных моментов основного и возбужденного состояний. Это различие в электронных дипольных моментах обусловлено разным электронным распределением в двух состояниях, и его можно считать отражением миграции заряда при переходе. Когда интеграл переходного момента обращается в нуль, переход становится запрещенным. [34]
Согласно так называемой концепции твердого состояния, ламеллы состоят из плотно упакованных, жестко ориентированных молекул, которые допускают миграцию зарядов, а не фотонов. По существу предполагается, что при каждом возбуждении происходит удаление электрона из поглощающей молекулы пигмента, после чего электрон ( так же, как и дырка, которая остается после удаления электрона) мигрирует к улавливающему центру. [35]
Значения RMS и особенно Кз и Kz незначительны при напряжении около 1 - 2 кВ, но с повышением напряжения они сильно возрастают. Отсюда ясно, что Kz действительно является константой скорости излучения, за счет которого происходит утечка зарядов в атмосферу-возможно, как через миграцию зарядов, ионизированных молекул воды, так и в результате притяжения заряженных частиц и молекул воды из окружающей среды. [36]
Энергия фотонов, поглощенная хлорофиллом, может передаваться по экситонному механизму. Однако до конца неясно, происходит ли разделение зарядов в том месте, где поглощаются дырки или электроны, или же экситон распадается в другом месте и затем происходит миграция зарядов. [37]
В этом случае принимается, что радикалы образуются в результате рекомбинации зарядов и по мере накопления радикалов ввиду того, что они обладают положительным сродством как к дырке, так и к электрону [3], все с большей вероятностью происходит миграция зарядов на радикалы, где затем они рекомбинируют без образования новых радикалов. [38]
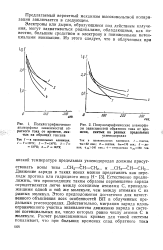 |
Полулогарифмические анаморфозы зависимостей обратного тока от времени, снятых на разных предельных. [39] |
Естественно предположить, что происходящее таким образом перемещение заряда осуществляется легче между соседними атомами С, принадлежащими одной и той же молекуле, чем между атомами С из разных молекул. Этого предположения достаточно для качественного объяснения всех особенностей ВП в облученных предельных углеводородах. Действительно, миграция заряда вдоль ограниченного участка эквивалентна движению заряда в системе потенциальных ям, число которых равно числу атомов С в молекуле. [40]
Одним из возможных механизмов передачи энергии в биологических системах является миграция заряда по системе пептидно-водородных связей в белковой молекуле. Настоящий обзор представляет собою попытку критически рассмотреть основные работы, посвященные этой проблеме. Следует подчеркнуть, что вопрос о возможности миграции заряда в белковых системах является дискуссионным и вызвал много противоречивых представлений, однако и в настоящее время он остается нерешенным. [41]
Указанный переход можно представить себе как колебания заряда вдоль оси г. Дипольный момент, связанный с этим движением, обусловливает появление яркой спектральной линии. В то же время, хотя переход Is - - 2s связан со значительной миграцией заряда, она происходит с сохранением его исходной сферической симметрии. Следовательно, в данном случае дипольный момент, связанный с осцилляциями из одного состояния в другое, отсутствует, свет не взаимодействует с атомом и такой переход не проявляется в спектре. [42]
 |
Сила F ( х, действующая на электрон, в зависимости от расстояния х [ Л. 6 - 24 ]. [43] |
Процесс осаждения длится до тех пор, пока не образуется устойчивый двойной электрический слой с полем, достаточным для того, чтобы сделать одинаковыми энергетические уровни положительных ионов обеих фаз; образование такого слоя связано с накапливанием положительных ионов в растворе вблизи поверхности электрода и электронов у внутренней поверхности металла. Когда электрохимические потенциалы ионов в металле и в растворе становятся одинаковыми, наступает равновесное состояние. Описанный процесс аналогичен рассмотренному ранее для двух находящихя в контакте металлов, с той лишь разницей, что миграция зарядов осуществляется при посредстве положительных ионов, а не электронов; энергетический уровень иона в растворе в значительной мере определяется энергией гидратации. [44]
В плане обсуждаемой проблемы интересно рассмотреть некоторые работы, посвященные электронному парамагнитному резонансу в облученных белках. Изучению ЭПР белков и родственных им веществ посвящено очень много работ отечественных и зарубежных авторов, и в рамках настоящего обзора невозможно и нецелесообразно их излагать. Остановимся лишь на некоторых из них, которые, по нашему мнению, являются важными для выяснения возможности миграции заряда в белке. [45]
Страницы: 1 2 3 4