Молекулы-акцептор
Cтраница 1
Молекулы-акцепторы должны диффундировать сквозь электролит; их попадание на металлическую поверхность будет беспорядочным, так же беспорядочным будет и удаление ионов металла. [1]
Рассмотрим атом р молекулы-акцептора, который наиболее близко соприкасается с молекулой-донором. [2]
Как было нами показано [2], для наблюдения этого эффекта необходимо, чтобы триплетный уровень молекулы-донора энергии был выше триплетного уровня молекулы-акцептора. Сенсибилизованную фосфоресценцию удобнее наблюдать в том случае, когда флуоресцентный уровень молекулы-донора энергии лежит ниже флуоресцирующего уровня молекулы-акцептора. Это дает возможность возбуждать избирательно молекулы-донора энергии, совершенно не затрагивая акцептор. [3]
Гинзбург [9] обнаружил, что при переходе от О к S в эфирах имеет место падение прочности водородной связи с о-хлорфено-лом, хотя ионизационный потенциал молекулы-акцептора при таком переходе тоже уменьшается. Прочность же комплексов с переносом заряда, образуемых подобными молекулами с фиксированным акцептором электрона, в таких рядах возрастает. [4]
Таким образом, изменение в распределении заряда при образовании связей В-N, которое, согласно расчетам, имеет преимущественно сг-ха-рактер, находит отражение в наблюдаемых ПИ остовных электронов атомов основания, а также молекулы-акцептора. ПИ B ( ls) отражает степень т-дони-рования и, следовательно, прочность связи В-N. Например, теплоты образования C5H5NBF3 ( 50 6 ккал / моль) [17] и C2H5NH2BF3 больше, чем для CH3CNBF3 ( 26 5 ккал / моль) [12], что согласуется со значениями ПИ C ( ls), из которых наблюдаемое для CH3CNBF3 является наибольшим ( 195 7 эВ), а для C5H5NBF3 - наименьшим ( 194 5 эВ) среди исследованных соединений. Результаты расчетов неэмпирическим методом ССП и полуэмпирическим методом ЧПДП согласуются с этой закономерностью в устойчивости комплексов. [5]
Причину, по которой некоторые атомы легко увеличивают свои координационные числа с трех до четырех, реже-с двух до четырех, можно понять, если рассмотреть полярный характер дативной связи. Как молекулы-доноры, так и молекулы-акцепторы электрически нейтральны. [6]
Есть другое доказательство того, что в обмене водорода с дейтерием могут играть решающую роль сильные электронодонорные и электроноакцеп-торные центры. Комплексы, содержащие как сильные, так и слабые молекулы-акцепторы, значительно менее эффективны. ЭДА-комплексы магния проявляют максимальную активность в обмене водорода на дейтерий, которая наблюдается в области более сильного сродства к электрону ( 1 2 - 1 6 эе), чем у комплексов натрия. [7]
Эндотермическая ( ДЯ13 кДж / моль) реакция должна быть примерно в 150 раз медленнее, чем процесс, для которого энергия начального и конечного состояний равны, при таком же значении энергии активации. Оказывается, что энергия, освобождающаяся при изменении геометрии молекулы-акцептора, переходящей из основного в триплетные состояния, может добавляться к общей энергии возбуждения: временной масштаб бимолекулярной реакции вполне достаточен для осуществления соответствующих скелетных смещений молекулы. Для акцепторных молекул, имеющих сильно разнесенные полосы ( 0 0) в случае процессов поглощения и испускания, такое поведение наиболее вероятно, что и подтверждается экспериментально. В тех ситуациях, когда геометрические изменения не обеспечивают достаточно энергии, скорость переноса быстро падает, как только процесс становится более эндотермичным. При использовании jjwc - стильбена в качестве акцептора скорость начинает быстро уменьшаться при значениях А. [8]
Таким образом, цикл трикарбоновых кислот не только участвует в конечном окислении питательных веществ, но служит также важным распределителем, поставляющим исходные соединения для синтеза основных структурных единиц ( строительных блоков) клетки. Если бы указанные кислоты постоянно выводились из цикла, то регенерации молекулы-акцептора не происходило бы и цикл был бы нарушен. Так называемые анаплеротические последовательности реакций обеспечивают поступление в цикл трикарбоновых кислот все новых количеств промежуточных соединений взамен израсходованных для биосинтеза. Эти анаплеротические последовательности имеют особо важное значение для тех организмов, которые растут за счет простых одно - или двух-углеродных соединений или других субстратов, разлагающихся на такие же простые соединения. [9]
Как было нами показано [2], для наблюдения этого эффекта необходимо, чтобы триплетный уровень молекулы-донора энергии был выше триплетного уровня молекулы-акцептора. Сенсибилизованную фосфоресценцию удобнее наблюдать в том случае, когда флуоресцентный уровень молекулы-донора энергии лежит ниже флуоресцирующего уровня молекулы-акцептора. Это дает возможность возбуждать избирательно молекулы-донора энергии, совершенно не затрагивая акцептор. [10]
Фотоок неленив - реакции окисления молекул вещества под действием света. Поглотив квант света, фотовозбужденные молекулы ( синглетновоз-бужденные, триплетные) вступают во взаимодействие с окислителями ( катионы, молекулы-акцепторы, кислород), образуя первично-окисленный продукт в результате переноса электрона на акцептор. [11]
АТР; их называют низкоэнергетическими соединениями. Под действием специфичных киназ фосфатные группы от сверхвысокоэнергетических фосфатов, образующихся в процессе катаболизма, могут переноситься на ADP, в результате чего синтезируется АТР. Другие специфичные киназы катализируют перенос концевой фосфатной группы АТР на молекулы-акцепторы, которые превращаются при этом в фосфо-рилированные ( низкоэнергетические) соединения и оказываются благодаря этому активированными; в таком виде они могут вступать в реакции биосинтеза. Следовательно, АТР в процессе метаболизма играет роль универсального промежуточного продукта-переносчика фосфатных групп. АТР также поставляет энергию для функционирования сократительных актиновых и миозиновых нитей скелетных мышц. За счет этой энергии осуществляется скольжение нитей друг относительно друга и как результат этого-сокращение мышцы; при этом АТР гидролизуется до ADP и фосфата. Креа-тинфосфат служит резервуаром высокоэнергетических фосфатных групп в мышечных и нервных клетках. Под действием креатинкиназы он может передавать свои фосфатные группы молекулам ADP. Химическую энергию для мембранных АТРаз тоже поставляет АТР; это дает им возможность переносить ионы Н и некоторые другие катионы через мембраны против градиента концентрации. [12]
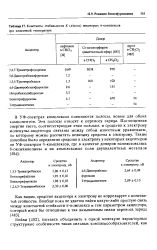 |
Константы стабильности К ( л / моль некоторых тт-комплексов при комнатной температуре. [13] |
В УФ-спектрах комплексов появляются полосы, новые для обоих компонентов. Поглощенная энергия света, соответствующая этим полосам, и сродство к электрону молекулы-акцептора связаны между собой известным уравнением, по которому можно рассчитать величину сродства к электрону. [14]
Реакция Михаэля обычно катализуется основанием - Все стадии обратимы; как правило, применяют протонные растворителя, особенно спирты, которые протонируюг промежуточный анионный аддукт и образуют продукт реакции. Реакция может контролироваться термодинамически или кинетически в зависимое. Наиболее обычны реакции с - такими нуклеофилами, как енолягы малоновых эфиров или 3-кегоэфиров, стабилизованных двумя электро-ноакцепторными заместителями. Получены также хорошие выходы продуктов алкилирования простых кетонов и нитроалканов, Енамнпы также используют в качестве нуклеофилов в реакции Михаэля. Разнообразные олефнны, в которых одна или несколько эдектрошаакцепторных групп сопряжены с двойной связью, применяют в качестве молекулы-акцептора. В ролы электрофилъшй частицы могут применяться также ацетилены. [15]
Страницы: 1 2