Муреин
Cтраница 2
Единственным возражением против этого может быть разница в составе клеточной стенки: у сине-зеленых водорослей, так же как и у бактерий, в клеточной стенке имеется муреин. В целом по составу и строению клеточной стенки, а также по реакциям, благодаря которым идет синтез ее веществ, прокариоты существенно отличаются и от животных, и от остальных растений. [16]
 |
Пакеты клеток Sarcina ventriculi, выросшие в питательной среде. ( Снимок в светлом поле, 750 х. фото D. Glaus. [17] |
Что касается биосинтеза экзополисахаридов, то следует отметить два момента: 1) декстраны и леваны синтезируются внеклеточными ферментами из дисахаридов; 2) состав большинства экзополисахаридов независим от используемого субстрата-путь их биосинтеза сходен с механизмом построения муреина или липополисахаридов. Активированные уридинтрифосфатом сахара ( UTP-caxapa) присоединяются к липиду-переносчику, комбинируются, образуя видоспецифические гомо - или гетерополимерные структурные компоненты будущего полисахарида, и выводятся из протопласта в наружные слои клеточной стенки, где связываются в макромолекулярный экзополисахарид. [18]
 |
Влияние терыореагентной обработки на скорость осаждения активного ила. [19] |
Полисахариды клеточных стенок условно разделяют на структурные и структурно-метаболические. Первые, например муреин, хитин, глюканы, образуют нерастворимый жесткий каркас, а вторые - достаточно растворимы. [20]
Входит в состав белков ( в фиброине шелка на его долю приходится до 40 %); в плазме крови содержится в свободном состоянии. В бактериальных клеточных стенках в составе муреина присутствуют L - и D-формы А. [21]
Общим у этих антибиотиков является наличие Д - лактамиого кольца. Механизм действия их состоит в торможении одной из стадий формирования муреина - пептидогликана, формирующего клеточную стенку бактерий. [22]
Во время инициации споры теряют до 30 % массы сухих клеток. Эксудат, в основном, состоит из ДПК, продуктов гидролиза муреина кортекса, пептидов, небольших белков, нуклеотидов РНК, дивалентных катионов. Споры обводняются, теряют свою рефрактерность, термостабильность и устойчивость к действию повреждающих агентов. Вследствие набухания споры увеличиваются в объеме чуть ли не вдвое. В связи с разрушением кортекса барьер проницаемости полностью снимается. Синтез макромолекул все еще репрессирован. [23]
Поперечные связи представлены пептидными мостиками. Вследствие разницы в химической структуре и, следовательно, в составе ферментов путей биосинтеза образование псевдомуреина не подвержено действию таких типичных для подавления синтеза муреина антибиотиков, как D-циклосерин, ванкомицин или пенициллин. [25]
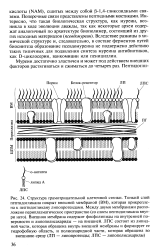
По сравнению с ЦПМ внешняя мембрана ( ВМ) является сильно асимметричным липид-ным бислоем, в котором внутренний слой состоит из фосфоли-пидов, а внешний - из липополисахаридов. ВМ и муреиновый слой соединены липопротеином, который своим N-кон-цом с замещенными жирными кислотами погружен во внешнюю мембрану, а его карбоксильный конец связан с муреином. ВМ содержит белки-порины, формирующие поры. [26]

Пептидогликан - структурный полимер, входящий в состав клеточных оболочек большинства прокари-отических микроорганизмов. Он состоит из линейных молекул гли-кана, соединенных между собой поперечно пептидами, содержащими 3 - 6 аминокислотных остатков. Гликан муреина образован остатками М - ацетил - М - глюкозамина и N-ацетилмурамовой кислоты, соединенных 3 - 1 4-связями. Незанятые в пептидных связях карбоксильные и аминогруппы аминокислот могут оставаться свободными или образовывать поперечные связи между пептидными мостиками. Аминокислотный состав и строение пептидов муреина имеют большое значение при идентификации грамположительных микроорганизмов. [28]
Наружная мембрана плотно прилегает к муреиновому слою и связана с ним липопротеинами. Муреиновый слой, видимо, свободно проницаем для различных веществ. Промежуток между муреином и плазматической мембраной называют периплазматическим пространством. В нем находятся белки, в том числе деполимеразы ( протеиназы, ну-клеазы, например рестрикционный фермент EcoRI), периферические белки плазматической мембраны и так называемые связующие белки. Последние участвуют в переносе некоторых субстратов в цитоплазму и служат рецепторами хемотаксических стимулов. [29]
Наружная мембрана плотно прилегает к муреиновому слою и связана с ним липопротеинами. Муреиновый слой, видимо, свободно проницаем для различных веществ. Промежуток между муреином и плазматической мембраной называют периплазматическим пространством. В нем находятся белки, в том числе деполимеразы ( протеи-назы, нуклеазы), периферические белки плазматической мембраны и так называемые связующие белки. Последние участвуют в переносе некоторых субстратов в цитоплазму и служат рецепторами хемотаксических стимулов. Периплазматическое пространство, по всей вероятности, играет также роль в осморегуляции. [30]
Страницы: 1 2 3