Протонный насос
Cтраница 2
Это можно объяснить тем, что протоны, выкачиваются из них, как это было описано выше. Присутствие протонного насоса объясняет и высокую концентрацию АТФ, отмечаемую в ситовидных трубках. [16]
Необходимо отметить, что натриевые насосы как системы активного транспорта характерны для структурных мембран клетки, первыми принимающими на себя воздействие внешней среды и не требующими для функционирования высокого электрического сопротивления. В этом случае действуют протонные насосы, которые служат главными узлами механизма сопряжения процессов окисления и фосфорилирования при генерации мембранного потенциала дыхательной цепью и АТФ-азой. [17]
По-видимому, теория Митчелла ( § 13.3) применима к этой системе. Имеются данные, указывающие на то, что зрительный родопсин также работает как фотогенератор, выполняя роль протонного насоса. [18]
 |
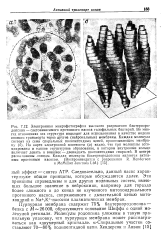
Электронная микрофотография нервно-мышечного синапса лягушки в момент слияния синаптическо-го пузырька с пре-синаптической мембраной и высвобождения медиатора. [19] |
У целого ряда бактерий имеются только два кольца в бактериальном моторе: 5-кольцо, прикрепленпое к стенке, и отстоящее от него на 3 5 нм М - кольцо, погруженное в клеточную мембрану и скрепленное со стержнем мотора; кольца состоят из 15 - 16 секторов. В живой бактериальной клетке часть энергии, получаемой ею при фотосинтезе и питании, используется для приведения в действие протонного насоса, выкачивающего протоны во внеш. Предполагают, что возникающий в результате этой разности концентраций поток протонов, проходящий между S - и Af-кольцами, приводит в действие бактериальный мотор; при этом ротор мотора совершает один оборот относительно статора при прохождении примерно 300 протонов. [20]
В недавних работах были проведены тщательные определения числа протонов, числа зарядов и числа молекул АТФ, приходящихся на каждый из трех центров сопряжения в ЦПЭ ( ср. С помощью различных доноров и акцепторов электронов были получены результаты, схематически показанные на рис. 13.9. Каждый центр сопряжения имеет протонный насос, разделяющий 4 заряда ( 4 протона выходят из цитоплазмы в мат-рикс) - отношение Hf / центр 4 во всех трех центрах. [21]
Следовательно, данный насос характеризуют общие принципы, которые обсуждаются далее. Эти принципы справедливы и для других модельных систем, имеющих большое значение в нейрохимии, например для гораздо более сложного и до конца не изученного митохондриального протонного насоса, сопряженного с дыхательной цепью митохондрий и Na K - насосом плазматических мембран. [23]
Группы из семи ос-спиральных столбов пронизывают мембрану и образуют молекулу размером 2 5 - 3 5 - 4 5 нм, причем наиболее протяженный участок располагается перпендикулярно поверхности мембраны. Ретиналь, по-видимому, связан с боковой цепью лизина и располагается между тремя внутренними и четырьмя внешними спиралями белковой молекулы. Каждая молекула бактериородопси-на представляет собой активный протонный насос. Функциональная роль тримера, обнаруженного методом электронной микроскопии, неизвестна. Модель Хендерсона и Анвина подтверждает основную концепцию: система ионного транспорта состоит из интегральных ( пронизывающих толщу мембраны) спирально уложенных полипептидных цепей. Поскольку аминокислотная последовательность бактериородопсина известна, в основном информация касается структуры этого ионного насоса. Как и в случае родопсина сетчатки, многое известно о фотохимии световой активации. Возможно, этот биохимически охарактеризованный протонный насос галофильных бактерий окажется первой системой с полностью выясненным функциональным механизмом. [24]
На первый взгляд энергозависимый синтез АТР, по-видимому, нельзя рассматривать как нейрохимическую проблему, но между передачей сигнала и энергетическим сопряжением существует некоторое сходство. Оба этих процесса имеют много общего и осуществляются с помощью белков, встроенных в липидные мембраны. Их взаимосвязь четко прослеживается при обсуждении фотозависимого протонного насоса у галофиль-ных бактерий ( с. Бактериальный рецептор, аналогичный рецепторам нейрона ( гл. Следовательно, энергия света внешнего сигнала обеспечивает внутриклеточный синтез АТР. Изучение бактерио-родопсина и механизма сопряжения фоторецепции, а также энергозависимого транспорта протонов ( и наконец, синтеза АТР) представляет особый интерес при исследовании нейро-рецептора. [25]

Ее цвет обусловлен наличием в ней бактериородопсина - пигмента, сходного с родопсином который содержится в зрительных клетках животных. Благодаря этому пигменту на свету создается протонный градиент между наружной и внутренней сторонами мембраны. Пурпурная мембрана выполняет, таким образом, функцию протонного насоса, приводимого в действие светом, и это ведет к возникновению электрохимического мембранного потенциала. Уравнивание зарядов может сопровождаться синтезом АТР: пурпурная мембрана делает возможным особый вид фосфорилирования. [27]
Протонный насос в отличие от других АТРаз синтезирует АТР благодаря наличию градиента протонов. Данная система выделена из митохондриальной мембраны, частично охарактеризована биохимическими методами и путем анализа реконструированных систем. Методом электронной микроскопии высокого разрешения определена трехмерная структура светоза-висимого протонного насоса галофильных бактерий. Все эти данные подтверждают ряд выдвинутых ранее гипотез о том, что такие транспортные системы состоят из а-спиральных полипептидных цепей, пронизывающих мембрану. [28]
Обратимость реакции АТР-синтазы имеет чрезвычайно важное значение для клетки. Например, образовавшийся в результате субстратного фосфорилирования АТР может быть использован АТР-синтазой для создания протонного градиента. Таким образом, этот фермент может также играть роль протонного насоса. Благодаря обратимости процессов, протекающих при этом в плазматической мембране, протонный градиент и АТР могут как бы взаимно превращаться друг в друга. [29]
Как полагают, быстрые реакции на гормон во всех случаях связаны с изменением проницаемости плазматической мембраны. Ауксин оказывает и более длительное действие на такие растущие клетки-оно связано с изменениями в транскрипции генов и с синтезом и секрецией нового материала для упрочнения клеточной стенки. Эти эффекты уже, по-видимому, не зависят от активации протонного насоса, и лежащие в их основе механизмы неизвестны. [30]
Страницы: 1 2 3