Натрий-калиевый насос
Cтраница 2
Неравнозначный перенос заряженных ионов ( частиц) через мембрану вызывает ее поляризацию: появление снаружи и - изнутри, поэтому натрий-калиевый насос называют электрогенным. Создаваемый градиент Na используется для вторичного активного транспорта глюкозы в клетки. Например, рассмотрим всасывание глюкозы из просвета кишечника: 1) переносчик глюкозы обеспечивает транспорт глюкозы в цитозоль энтероцита за счет входа в клетку ионов Na под действием электрохимического градиента ( концентрация Na: высокая - в просвете кишечника и низкая - в цитозоле клеток); 2) глюкоза из клетки переходит во внеклеточную жидкость по механизму облегченной диффузии. Скорость переноса глюкозы зависит от величины градиента Na: она увеличивается при повышении разности концентраций Na в просвете кишечника и цитозоле энтероцита; 3) Na, К - АТФаза поддерживает эту разность концентраций Na за счет его откачки в межклеточное пространство в обмен на К с затратой АТФ. Согласованный перенос двух веществ называют котранспортом: при симпорте имеет место перенос обоих веществ в одном направлении, а при антипорте - в противоположных направлениях. [16]
При решении вопроса о том, какие именно ионы принимают участие в генерации пейсмекерных потенциалов, первой гипотезой, требующей рассмотрения, является представление о натрий-калиевом насосе. В обычных условиях сбалансированного переноса ионов натрия из клетки, а ионов калия в клетку этот активный транспорт электронейтрален. Преобладание вывода ионов натрия создает гиперполяризационный электрогенный эффект. Преобладание активного ввода ионов калия создает де-поляризационный эффект. Достаточно допустить, что электронейтральный в целом натрий-калиевый насос начинает работать со смещением по фазе для активного транспорта натрия и калия, как активный транспорт становится электрогенным. Фаза деполяризации соответствует преобладанию переноса в клетку калия, а фаза поляризации - выводу натрия. [17]
Он обеспечивается за счет фермента аденозинтрифосфатазы мембран кардиомиоцитов, который, в свою очередь, регулируется тонкими биохимическими реакциями. Натрий-калиевый насос поддерживает на стабильном уровне высокие значения потенциала покоя, что имеет решающее значение в обеспечении функции сократимости миокарда. Для его поддержания необходимо, чтобы выходящий из кардиомио-цита калиевый ток был равен по величине и противоположен по направлению натриевому току. Превышение входящего тока над выходящим предопределяет величину потенциала действия и скорость его нарастания, чем регулируются потоки кальция и, в итоге, процесс актомиозинового сопряжения. [18]
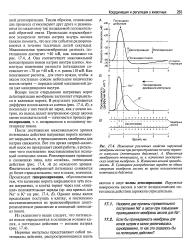
После достижения максимального уровня потенциала действия натриевые ворота начинают закрываться, и проницаемость мембраны для натрия снижается. Все это время натрий-калиевый насос не прекращает своей работы, в результате чего постепенно восстанавливается исходный потенциал покоя. Реполяризация приводит к снижению пика, или спайка, потенциала действия ( рис. 17.4, А) до исходного уровня. Фактически мембранный потенциал снижается до более отрицательного, чем в покое, значения. Происходит гиперполяризация, обусловленная тем, что калиевые ворота закрываются чуть позже натриевых, и клетка теряет через них лишние положительные заряды ( ср. Однако ионы калия продолжают поступать в клетку, и постепенно восстанавливаются их трансмембранное электрохимическое равновесие и исходный потенциал покоя. [20]
Плохая ионная пропускная способность и большое электрическое сопротивление мембраны способствуют поддержанию такого энергизованного состояния. Цианистый калий подавляет действие натрий-калиевого насоса. Поэтому отравление нервных волокон цианистым калием приводит к постепенному выравниванию концентраций ионов Na и К 1 внутри и вые аксона. [21]
В нервных и мышечных клетках натрий-калиевый насос обеспечивает возникновение в плазматической мембране разности потенциалов, называемой потенциалом покоя ( о проведении нервных импульсов см. гл. [22]
Адаптация при инъекции анионов, по-видимому, состоит в том, что включается активный вывод хлора, создавая эффект деполяризации, компенсирующий гиперполяризационный сдвиг и ответственный за деполяриза-ционное последействие. Другая возможность заключается в том, что инъекция анионов смещает баланс в натрий-калиевом насосе так, что начинает преобладать ввод калия, в результате чего возникает электрогенный эффект в виде деполяризации, ограничивающей гиперполяризацию, вызванную введением анионов. [23]
На рис. 13.5 показана принципиальная схема натрий-калиевого насоса, а на рис. 13.3 дана схема генератора Ван-де - Граафа, в котором при коронном разряде заряды переносятся лентой и накапливаются на полусфере, расположенной на изоляционных подставках. В своей основе эти два ЭП близки друг к другу: в натрий-калиевом насосе заряды переносятся белковой молекулой, которая вращается, а в генераторе Ван-де - Граафа заряды переносятся бесконечной лентой. Как в том, так и другом случае заряды переносятся медленно и токи небольшие. [24]
Одной из наиболее интересных проблем биохимии является превращение химической энергии в механическую, что составляет основу таких биологических процессов, как мышечное сокращение, транспорт веществ между телом нейрона и синапсами, а также активный транспорт ионов и молекул через клеточную мембрану. Было подсчитано, что в состоянии покоя - 30 % энергии дыхания используется на работу натрий-калиевого насоса. [25]
Калий в отличие от натрия может не только реабсорбироваться, но и секретироваться. При секреции калий из межклеточной жидкости поступает через базальную плазматическую мембрану в клетку канальца за счет работы натрий-калиевого насоса, а затем выделяется в просвет нефрона через апикальную клеточную мембрану пассивно. Секреция, как и реабсорбция, является активным процессом, связанным с функцией клеток канальцев. Механизмы секреции те же, что и механизмы реабсорбции, но только все процессы протекают в обратном направлении-от крови к канальцу. [26]
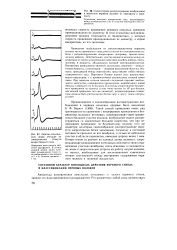 |
Сальтаторное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату.| Ответы седалищного нерва лягушки на электрические стимулы возрастающей силы. [27] |
Такой способ проведения имеет ряд преимуществ по сравнению с непрерывным проведением в безмякотных волокнах: во-первых, перепрыгивая через сравнительно большие участки волокна, возбуждение может распространяться со значительно большой скоростью, чем при непрерывном проведении: по безмякотному волокну того же диаметра; во-вторых скачкообразное распространение является энергетически более экономным, поскольку в состояние активности приходит не вся мембрана, а только ее небольшие участки в области перехватов, имеющие ширину менее 1 мкм. Потери ионов ( в расчете на единицу длины волокна), сопровождающие возникновение потенциала действия в таких ограниченных участках мембраны, очень невелики, а следовательно, малы и энергетические затраты на работу натрий-калиевого насоса, необходимые для восстановления измененных ионных соотношений между внутренним содержимым нервного волокна и тканевой жидкостью. [28]
 |
Повышение потребления кислорода нервом в зависимости от частоты стимуляции. [29] |
Нерв в состоянии покоя потребляет кислород и выделяет углекислоту. Если один участок нерва поместить в атмосферу азота, а другой оставить в воздухе, то уже через несколько минут можно обнаружить, что поверхность нерва, лишенная кислорода, приобретает электроотрицательный заряд. Объясняется это тем, что в бескислородной среде вследствие изменения обменных процессов нарушается работа натрий-калиевого насоса, в результате чего происходит прогрессирующая деполяризация мембраны нервных волокон. Примерно через 1 / 2 ч потенциал покоя падает настолько, что проведение нервных импульсов на этом участке нерва полностью прекращается. При возвращении нерва в атмосферу кислорода потенциал покоя быстро восстанавливается и даже на некоторое время становится больше исходной величины. Одновременно восстанавливается и проведение возбуждения. [30]
Страницы: 1 2 3