Образование - катехин
Cтраница 1
Образование катехинов ( измеряемое по реакции с ванилиновым реактивом) в коре трех видов ив - Salix elegantissima, S. Поскольку в этом случае побеги не имели листьев, а несли лишь нераспустившиеся лм - стовые почки, очевидно, что катехины синтезировались из каких-то запасных веществ, а не за счет продуктов фотосинтеза. Позднее Курсанов и Крюкова ( 1950) показали. Сходные результаты были получены Дурмишидзе ( 1955) для взрослых листьев виноградной лозы сорта Саперави, хотя в этом случае образование кате-хиновых дубильных веществ было относительно более слабым, чем в листьях чая. [1]
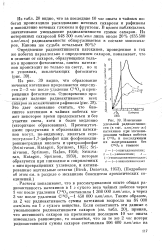 |
Изменение удельной радиоактивности ( в тыс. имп / мин-мг катехинов при экспонировании чайных побегов в С1402 и последующем их выдерживании без CuOj в темноте. [2] |
Из рис. 39 видно, что образование меченых катехинов продолжается еще спустя 2 - 3 час после удаления С1402 и прекращения фотосинтеза. [3]
Из табл. 21 видно, что все испытанные предшественники использовались для образования катехинов ( этилацетатная фракция), хотя и в различной степени. Через 6 час после введения наибольшее включение метки в состав катехинов ( 12 8 %) происходит в опыте с шикимовой кислотой. Слабее включается во фракцию катехинов ацетат ( 7 3 %) и еще более слабо фруктоза, глюкоза и сахароза. [4]
Как можно видеть из данных табл. 3, процессы роста и образования катехинов наиболее резко тормозятся в отсутствии азота в питательной смеси. Дополнительное внесение / 2 дозы NPK вместе с резким увеличением роста и образования органических веществ приводит к усилению синтеза катехинов. [5]
Раздельный анализ нормальных и этиолированных проростков позволял выяснить роль световой энергии в образовании катехинов на ранних стадиях развития чайного растения. [6]
В 1953 г. одномерная хроматография на бумаге была использована ( Запрометов, 1953) для изучения образования катехинов при прорастании семян чая. В качестве растворителя была подобрана однофазная смесь: к-бутанол СН3СООН Н2О ( 40: 12: 29 по объему), дающая лучшее разделение катехинов по сравнению с ранее применявшимися для фенольных соединений растворителями. Поскольку на начальных стадиях прорастания семян в них образуется лишь ( -) - эпикатехин, то его количество было можно определить по реакции с ванилином. [7]
Из табл. 13 видно, что отделение листьев и почки от стебля приводит к заметному снижению скорости образования всех катехинов. В среднем скорость образования катехинов снижается на 35 %, что, возможно, частично зависит от менее благоприятного расположения отделенных листьев и фотосинтетической камере. [8]
В отличие от приведенных данных, изучение биосинтеза катехинов в побегах чайного растения ( Запрометов, 1954) показало, что уже через 3 час световой экспозиции в атмосфере с С1402 достигается высокая радиоактивность катехинов. Поэтому была предпринята попытка изучить скорость образования катехинов, пользуясь еще более короткими экспозициями. [9]
Хроматограмма наземных частей этиолированных проростков отличается от хроматограммы нормальных проростков лишь меньшим содержанием ( -) - эпигаллокатехина ( более слабая окраска и меньшие размеры пятна) и более коротким хвостом. На основании этих данных следует, что образование катехинов в проростках чайного растения может происходить и без участия световой энергии за счет имеющихся запасных продуктов. В то же время освещение проростков благоприятствует синтезу в них ( -) - эпигал-локатехина. [10]
Приведенные данные свидетельствуют о способности листьев чайного растения и виноградной лозы к синтезу катехинов. Однако размеры этой биологической способности, значение листьев в общем полифенольном балансе растения, а также последовательность образования катехинов могли быть изучены только с применением изотопной методики. [11]
Поскольку в данном случае возможность оттока катехинов из взрослых листьев была исключена, следует признать что основным местом образования катехинов в чайном растении являются молодые побеги. [12]
Еще большее значение имеют гидрированные производные окси-флавонов, так называемые антоцианидины, являющиеся солями бензо-пирилия. Так, при восстановлении кверцетина может быть получена бензопирилиевая соль ( цианидин); его дальнейшее восстановление приводит к образованию катехина, находящегося в дубильных экстрактах и в чае. [13]
Еще большее значение имеют гидрированные производные окси-флавонов, так называемые антоцианидины, являющиеся солями бен-зопирилия. Так, при восстановлении кверцетина может быть получена бензопирилиевая соль ( цианидин); его дальнейшее восстановление приводит к образованию катехина, находящегося в дубильных экстрактах и в чае. [14]
Суммируя изложенное, можно сделать вывод, что основным местом синтеза катехинов являются молодые побеги чайного растения, где катехины образуются из продуктов фотосинтетической ассимиляции углекислоты. Поскольку растения с отделенными корнями осуществляют синтез катехинов примерно в тех же размерах, что и целые растения, следует признать, что образование катехинов ( а следовательно, и более уплотненных дубильных веществ катехинового типа) в листьях и молодых побегах не имеет непосредственной связи с деятельностью корневой системы. [15]
Страницы: 1 2