Однонитевая
Cтраница 1
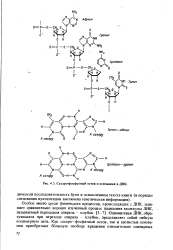 |
Сахаро-фосфатный остов и основания в ДНК. [1] |
Однонитевая ДНК, образующаяся при переходе спираль - клубок, представляет собой гибкую полимерную цепь. [2]
Лигандом служила однонитевая тимусная ДНК. На колонку, уравновешенную буфером, содержащим 0 15 М NaCl, с очень малой скоростью вносили осветленный ( 12 000 об / мин, 30 мин) лизат клеток. [3]
РНК, чаще всего однонитевая. [4]
 |
Задний фонарь. [5] |
На автомобиле Москвич-412 задний фонарь четырехламповый: одна - указатель поворота, вторая - габаритный фонарь, третья - сигнал стоп и четвертая - освещение при движении задним ходом ( рис. 65, а), а на автомобиле ВАЗ-2101 - двухламповый ( рис. 65, б) с однонитевой и двухнитевой лампами: двух-нитевая лампа - указатель поворота и сигнал стоп, однонитевая - габаритный фонарь. [6]
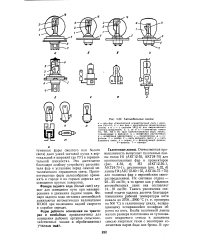 |
Автомобильные лампы. [7] |
Европейский асимметричный свет с цоколем Р45 / 41; б - для фар с европейским, симметричным спетом; ей г - с цоколем 2ФД - 42 для американского слетораспределения; д, е, ж и з - галогенные лампы Н4, НЗ, Н2 и HI ( для противотуманных фар); лампы, применяемые в фонарях: и - штифтовая двухннтевая; к - штифтовая однонитевая; л - пальчиковая; м - си-фятная: / - колба; 2 - нить дальнего света; 3 - нить ближнего света; 4 - экран; 5 - фиксирующий фланец; б-цоколь; 7 - выводы. [8]
Конечно, дело не ограничивается спиральными участками. Однонитевая ДНК склонна принимать весьма причудливую пространственную конфигурацию. При этом замкнутость цепи в кольцо неизбежно вызывает напряжения, которые могли бы исчезнуть, будь кольцо разомкнуто. [9]
Колонку промывают 5 - 10 объемами того же ( подогретого) буфера; в первых порциях элюата из колонки выходит однонитевая ДНК. Затем, не снижая температуры, увеличивают концентрацию элюирующего фосфатного буфера до 0 4 - 0 5 М; при этом с оксиапатита десорбируется двунитевая ДНК. Количество той и другой ДНК можно оценить по площади пиков оптической плотности с учетом гиперхромизма однонитевой ДНК или по радиоактивности. [10]
Совершенно иначе функционируют ретровирусы. С помощью этого фермента в зараженной клетке производится единственная однонитевая ДНК-копия вирусной РНК, которая при участии ферментов хозяина превращается в двунитевую ДНК. По ходу обратной транскрипции в качестве промежуточных образований возникают гибридные ДНК-РНК-структуры, одна цепь которых происходит из РНК вириона, а другая - из продукта обратной транскрипции. По мере разрушения РНК синтезированная однонитевая ДНК становится матрицей для формирования комплементарной ДНК-цепи. В дальнейшем новые молекулы вирусной РНК и вирусных белков, необходимые для образования новых вирусных частиц и развития инфекции, производятся с участием общих систем транскрипции и трансляции клеток хозяина. [11]
Вирус ВИЧ в составе зрелой внеклеточной частицы содержит геномную молекулу РНК. Однако развитие вирусной инфекции связано с этой РНК лишь в самой начальной фазе. По информации, содержащейся в этой РНК, воспроизводится молекула ДНК, сначала однонитевая. Затем на ней, как на матрице, воспроизводится комплементарная цепь и образующаяся двунитевая ДНК встраивается в геном инфицированной клетки. Именно эта ДНК далее управляет синтезом копий мРНК, необходимых для программирования синтеза вирусных белков, и самой РНК вириона, которая должна входить в состав новых вирусных частиц. Этот фермент катализирует гидролитическое расщепление РНК в составе гибридного дуплекса РНК-ДНК. Оказывается, все эти активности присущи самой обратной транскриптазе вируса ВИЧ-1. Однако самое удивительное состоит в том, что переключение активностей происходит строго упорядоченно, это не просто три параллельно работающих активных центра на одном ферменте. Рассмотрим ее более подробно. [12]
Однако в продукте реакции, кроме кДНК, синтезированной по матрице мРНК, неизбежно присутствует и вторичный продукт - ДНК, синтезированная уже по матрице самой кДНК, в том числе и двунитевые молекулы, состоящие из кДНК и вторичной ДНК. В этих условиях РНК и однонитевая ДНК не сорбируются. Остаточные дуплексы ДНК-ДНК в этих условиях не элюируются. [13]

У этого организма практически все виды гомологичной рекомбинации зависят от функционирования гена reck ( этот ген играет важную роль и в репарации повреждений ДНК; см. гл. Оказалось, что RecA-белок ( молекулярная масса около 40 кД) является ДНК-зависимой АТРазой, способной заменять одну из цепей дуплекса ДНК на гомологичную. Процесс поиска является активным и требует гидролиза АТР. Показано, что дуплекс ДНК в комплексе с RecA-белком и одноцепочечной ДНК заметно раскручен ( вне зависимости от того, гомологична однонитевая ДНК дуплексу или нет); по всей вероятности, это облегчает поиск гомологии. Другими словами, RecA-белок активно проводит однонитевую миграцию ветви. [15]
Страницы: 1 2