Биогенез
Cтраница 2
Задача изучения биогенеза этилена должна была бы упроститься, если бы удалось получить субклеточные фракции плода, способные осуществлять синтез этилена. Известно несколько сообщений, в которых утверждается, что такие препараты удалось получить и что удалось синтезировать этилен из специфических предшественников. Мей [69], однако, показал, что большинство этих работ ошибочно. Чандре и Спенсеру [30] все же удалось получить субклеточные фракции, которые действительно продуцировали этилен. [16]
Изучение механизма биогенеза стероидов, каротиноидов, каучуков, терпенов, боковых цепей витаминов К и Е, убихинонов указало на ступенчатое превращение исходного для всех этих соединений - ацетата в мевалоновую кислоту и распад ее 5-пирофосфата в пирофосфорный эфир изопентенилового спирта. Фермент изопентенил-пирофосфатизо-мераза катализирует превращение этого эфира в пирофосфат диметил-аллилового спирта. Конденсация двух пирофосфорных эфиров и последующие реакции представляют путь энзиматического образования изопреноидных соединений ( новых С-С - связей) без участия карбонильных групп. [17]
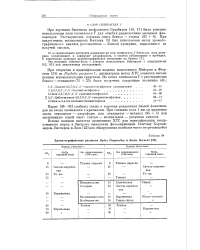 |
Хроматографические различия Radix Pimpinellae и Radix Heraclei. [18] |
При изучении биогенеза изофлавонов Гризбахом [16, 17] были успешно использованы слои силикагеля Г для очистки радиоактивно меченных фла-воноидов. При аналогичных исследованиях Биллека [5] был использован метод хромато-графического анализа ( растворитель - бензол) кумарина, выделенного из пахучей смолки. [19]
Данные по биогенезу мембран также не подтверждают концепцию строения мембраны из субъединиц. В этом случае сборка мембраны должна была бы быть одноэтапным процессом, в то время как в действительности сборка мембраны происходит в несколько этапов: вначале создается липидная основа мембраны, а далее присоединяется белковый компонент. [20]
Вопрос о биогенезе стероидов особенно интенсивно разрабатывался в последние годы в связи с применением меченых атомов. [21]
Помимо энергии, биогенез ДНК требует наличия специфических ферментов, катализирующих отдельные этапы синтеза, и множества белковых факторов, абсолютно необходимых для регулирования процесса репликации и проявления каталитической активности ферментов. [22]
Со стадии латирана биогенез касбеноидов направляется по трем путям. Так, в результате расщепления циклопропанового кольца и циклизации образуются вещества с углеродным скелетом мирсинана. [23]
Центральной проблемой теории биогенеза является вопрос: какая из концепций - чисто механическая, термодинамическая, статистическая или та, которая составляет содержание неравновесной статистической механики - позволяет предвидеть образование систем, хотя бы отчасти напоминающих живые. [24]
Несколько слов о биогенезе стероидов. [25]
Что касается взглядов на проблему биогенеза со стороны термодинамики, то сегодня подробно обсуждать их уже не имеет смысла. Пригожиным обобщенная термодинамика [11] снимает препятствия, воздвигнутые классической термодинамикой на пути самоорганизации материальных систем. [26]
Таким образом, происходит смена биогенезов на одной и той же территории ( суксеция) в результате природных и антропогенных факторов. [27]
Весьма интересны данные о путях биогенеза гризеофульвина и близких ему антибиотиков. Как известно, биохимическое построение углеродных цепей часто рассматривается как процесс последовательного соединения остатков уксусной кислоты, например, под влиянием коэн-зима А. Развитие этих взглядов в применении к биосинтезу природных фенольных соединений привело в 1953 г. к предположению, что аналогичным путем образуется и скелет гризеофульвина, причем гидрированное орсиновое кольцо антибиотика возникает в результате альдольной циклизации, а флороглюциновое - в результате внутримолекулярного С-ацшшрования. [28]
Как и в других случаях сесквитерпенового биогенеза, предшественником этих веществ выступает фарнезилпирофосфат. Пути же, ведущие от универсального прародителя к соединениям табл. 2, разнообразны. [29]
Более важны с точки зрения биогенеза реакции гидроксилирования и декарбоксилирования. Гидроксилированные в бензольном кольце производные триптофана образуются с помошью специфических ферментов-оксидаз. Особенно важен 5-гидрокситриптофан 6.376. Декарбоксилирование индольных а-аминокислот ведет к синтезу индолил-3 - этиламинов. [30]
Страницы: 1 2 3 4