Биосинтез - лизин
Cтраница 2
Большинство продуцентов лизина являются мезофильными микроорганизмами, имеющими оптимум роста, развития и биосинтеза лизина при температуре 28 - 30 С. Отклонения от этой температуры нежелательны. Повышение температуры на 5 - 7 С от оптимальной приводит к быстрому автолизу культуры, и в среде накапливается значительна меньше лизина. Понижение температуры на 4 - б С также невыгодно, так как культивирование резко затягивается на 12 - 20 ч, хотя выход ли-зкна при этом почти не падает. [16]
Так, превращение аспарагиновой кислоты в аспартилфосфат, катализируемое аспартаткиназой, является первой стадией биосинтеза лизина, метионина, треонина и изолейцина. Последний, как мы видели, синтезируется прямо из треонина, тогда как синтез лизина и метионина идет по другим путям. Треонин регулирует скорость своего синтеза благодаря способности ингибировать аспартаткина-зу. Однако если бы треонин полностью ингиби-ровал аспартаткиназу, то прекратился бы синтез и еще одной аминокислоты - лизина. [17]
Следует также учитывать, что и другие аминокислоты, не участвующие в обмене при биосинтезе лизина, могут оказывать влияние на рост и развитие микроорганизма. Их влияние учитывается каждый раз при работе с новым продуцентом - аминокислотный состав среды должен уточняться экспериментально. [18]
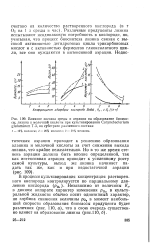 |
Влияние состава среды и аэрации на образование биомассы, лизина и молочной кислоты при культивировании Corynebacterium glutamicum T-3, на субстрате различного состава. [19] |
Различные продуценты лизина испытывают неодинаковую потребность в кислороде, но, учитывая, что процесс биосинтеза лизина связан с высокой активностью дегидрогеназ цикла трикарбоновых кислот и с активностью ферментов глиоксалатного цикла, все они нуждаются в интенсивной аэрации. [20]
Различные продуценты лизина испытывают неодинаковую потребность в кислороде, но, учитывая, что процесс биосинтеза лизина связан с высокой активностью дегидрогеназ цикла трикарбоновых кислот и с активностью ферментов гли-оксилатного цикла, все они нуждаются в интенсивной аэрации. [21]
В настоящее время отмечается большой дефицит кукурузного экстракта, используемого в качестве источника ростовых факторов при биосинтезе лизина. Необходимо изыскать заменитель экстракта, использование которого не ухудшило бы экономику производства лизина. [22]
Однако аминоадипиновый путь биосинтеза лизина не имеет места у промышленных продуцентов лизина, вернее у подавляющего большинства продуцентов лизина. Схема регуляции и биосинтеза лизина представлена ниже. [23]
 |
Регуляция биосинтеза лизина у бактерий. [24] |
Аспарагиновая кислота при биосинтезе лизина образуется у глутаматсинтезирующих бактерий за счет реакции переаминирования, которая осуществляется между щавелевоуксусной и глутаминовой кислотами. [25]
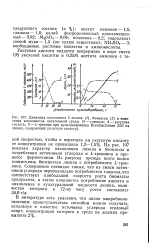
На рис. 107 показан характер накопления лизина и биомассы и потребления источников углерода и L-треонина в процессе ферментации. Из рисунка прежде всего видна взаимосвязь биосинтеза лизина с потреблением L-треонина. [27]
Условия культивирования определяются рН среды, степенью аэрации среды, длительностью и температурой ферментации, возрастом и дозой посевного материала. Все эти факторы оказывают существенное влияние на биосинтез лизина. [28]
D-Лизин используется для рост. Neurospora, у которых нарушены ранние стадии биосинтеза лизина; штаммы, у которых блокировано ближайшее к лизину звено биосинтетического процесса, не могут использовать D-лизин. [29]
Все продуценты лизина являются биотинзависимыми микроорганизмами. Если же снизить количество биотина до 1 - 2 мкг / л, биосинтез лизина снижается в 20 - 30 раз, но одновременно резко повышается биосинтез глутаминовой кислоты. [30]
Страницы: 1 2 3