Биосинтез - пиримидин
Cтраница 2
 |
Концентрация дезоксиуридина в тканях животных после общего рентгеновского облучения дозой 650 р. [16] |
Не касаясь различий в механизмах накопления тимидина и дезоксиуридина, на основании этих данных можно отметить, что наблюдавшееся повышение концентрации дезоксирибозидов, вероятно, не может быть безразличным для анаболизма пирими-динов. Действительно, как показали Калдарера и др., активность ключевого фермента биосинтеза пиримидинов аспартат-карбамилтрансферазы в печени крыс через 48 час. [17]
В живых организмах встречаются также и другие пуриновые и пирими-диновые основания, которые, однако, не входят в состав нуклеиновых кислот. К ним относятся: оротовая кислота, играющая роль промежуточного продукта при биосинтезе пиримидинов ( см. стр. С другой стороны, нуклеотиды этих соединений - инозиновая и ксантиловая кислоты - являются ключевыми промежуточными продуктами в биосинтезе пуринов ( см. стр. Замещенные окисленные пурины теофиллин, теобромин и кофеин входят в состав важных соединений растительного происхождения. [18]
При отрицательной обратной связи продукты ферментативных реакций снижают либо активность, либо синтез одного или нескольких взаимосвязанных ферментов. Например, адениловая и гуаниловая кислоты подавляют активность первого в цепи биосинтеза пуриновых оснований фермента - фосфорибозилпирофосфат - а ми до-трансферазы, а цитидиловая кислота регулирует биосинтез пиримидинов, аллостери-чески воздействуя на аспартат - карбамоилтрансферазу, катализирующую образование карбамоиласпартата из карбамоилфосфорной кислоты и аспартата. [19]
 |
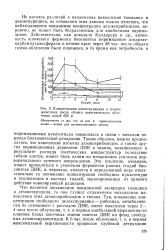
Биосинтез пиримидиновых нуклеотидов. [20] |
ИМФ из простых предшественников, которые, по-видимому, не связаны друг с другом. Что касается высших организмов, то несколько из этих ферментов образуют мультиферментный комплекс. Выживающие клетки преодолевают ингибирующее действие ФФА, синтезируют АТК-азу, причем больше примерно в 100 раз, чем нормальные клетки, увеличивается концентрация и карбамоил-синтетазы и дигидрооротазы. В то же время активность ферментов, катализирующих последующие реакции биосинтеза пиримидинов, практически не изменяется. [21]
Как правило, они блокируют некоторые стадии биосинтеза обычных пуриновых и пиримидиновых нуклеотидов. Обычно блокирование происходит после того, как сам ингибитор превращается в свой нуклео-тид. Так, 4-азаурацил ( иногда обозначаемый в литературе как 6-азаурацил) сначала превращается в свой нуклеозид, затем в 4-азауридинмонофосфат. Последний и оказывает ингибирующее действие на оротидин-5 - фосфатдекарбоксилазу, препятствуя тем самым биосинтезу пиримидинов [76, 77]; 5-фторурацил, оказавшийся сильным ингибитором роста некоторых опухолей, превращается сначала в свой рибонуклеотид, потом в дезоксирибо-нуклеотид и уже в такой форме [78] оказывает блокирующее действие, подавляя превращение дУМФ в дТМФ ( стр. [22]
 |
Механизмы, регулирующие биосинтез адениновых и гуаниновых нуклеотидов у.. coli по типу обратной связи. У других организмов эти биосинтетические пути регулируются иначе. [23] |
Обычные гшримидиновые нуклеотиды-это цитидин-5 - монофосфат ( СЫР), или цитидилат, и уриаин - У-монофосфат ( UMP), или уридилат; в их состав входят пиримидиновые основания-соответственно цитозин и урацил. Биосинтез пиримидиновых нуклеотидов ( рис. 22 - 19) несколько отличается от синтеза пуриновых нуклеотидов; в этом случае сначала образуется шестичленное пи-римидиновое кольцо, а затем к нему присоединяется рибозофосфат. Для этого процесса требуется карбамоилфосфат, играющий роль промежуточного продукта также и в цикле мочевины ( разд. Однако карбамоилфосфат, необходимый для синтеза мочевины, образуется в митохондриях при участии митохон-дриального фермента карбамоилфосфат-синтетазы I, а карбамоилфосфат, участвующий в биосинтезе пиримидинов, образуется в цитозоле под действием другой формы того же фермента - кар-бамоилфосфат-синтетазы II. Цитозоль-ный карбамоилфосфат взаимодействует с аспартатом, в результате чего получается N-карбамоиласпартат. Эта реакция катализируется аспартшп-транс-карбамоилазой, одним из наиболее тщательно изученных аллостерических ферментов ( дополнение 9 - 5); о ней мы еще будем говорить ниже. [24]
Синтез пуринового скелета начинается с конденсации глицина с амино-рибозо-5 - фосфатом. В процессе синтеза в качестве промежуточного продукта образуются риботиды с незаконченной пуриновой структурой. Процесс протекает так, что связь с рибозо-5 - фосфатом не нарушается в течение всего синтеза. Поэтому фактически образуются не свободные пуриновые осноняния. На ряде этапов этого синтеза в нем принимает участие АТФ. LB биосинтезе пиримидиновых оснований, точнее пиримидиновых нуклеотидов, важнейшую роль играет аспарагиновая кислота. Установлено, что в процессе биосинтеза пиримидинов из нее и карбамилфосфата ( стр. Оротовая кислота ( точнее, ее нуклеозид - оротидин-5 - фосфат) затем декарбоксклируется с образованием уридин-5 - - фосфата. [25]
Страницы: 1 2