Оогония
Cтраница 2
Очень часто спорангии и оогонии с антеридиями образуются на одном и том же растении одновременно. Первые зооспорангии располагаются по периферии колонии. При последующем симподиальиом ветвлении гиф они сдвигаются в сторону. Вторичные зооспорангии образуются симподиально. Зооспоры выходят так же, как и у моноблефариса. [16]
Страсбургер утверждал, что оогонии и антеридии циклоспоровых подобны тетраспо-рангиям диктиотовых водорослей, так как они развиваются на диплоидных слоевищах и первое деление ядра сопровождается мейозом. Но сами яйцеклетки и антерозоиды Страсбургер сравнивал с гаметами других водорослей, считая, что тетраспоры сразу, минуя стадию гаме-тофита, делятся и дают гаметы. К ю л и н вначале ( в 1916 - 1917 гг.) полагал, что цикл развития циклоспоровых возник в результате полного недоразвития гаметофита, а зооспоры ( споры) стали сразу функционировать как гаметы. Позднее, в 1937 г., Кюлин выдвинул теорию, согласно которой циклоспоровые возникли независимо от остальных бурых водорослей и у них никогда не было гаметофита, а их предки были всегда диплоидными. Из-за отсутствия убедительных данных, неоспоримо свидетельствующих в пользу какой-либо из этих теорий, ни одна из них не стала общепризнанной. [17]
Большинство Sapro-legniaceae гомоталличны: оогоний и оплодотворяющие их антеридии образуются на одном и том же вегетативном теле. Антеридии меньше по своим размерам и образуются на кончиках гиф. Зрелые антеридии ( по одному или несколько) прикрепляются к оогонию. Их оплодотворяющие трубки проникают к оосферам через стенку оогония. Когда одно из ядер антеридия достигнет соответствующего ядра яйцеклетки и сольется с ним, образуется диплоидное ядро зиготы. Оплодотворенная оосфера окружается толстой стенкой и превращается в ооспору. После длительного периода покоя ооспоры прорастают; при этом происходит редукционное деление. Цикл завершается образованием нового спорангия. [18]
В результате полового процесса - слияния оогония и антеридия - образуются ооспоры. Они распространяются внутри пораженной ткани и с отмершими растениями остаются после уборки урожая в почве или на ее поверхности, сохраняя жизнеспособность в течение двух-трех лет. [19]
При половом процессе на мицелии формируются оогонии и антеридии. Оогонии образуется в виде вздутия на конце сравнительно короткой ветви-ножки, отделяясь от нее перегородкой в основании. В оогонии формируется несколько круглых яйцеклеток. Антеридий представляет собой конечную клетку тонкой антеридиальной ветви. Он прикладывается плотно к оболочке оогония, в которой имеются более тонкие места - поры - и пускает внутрь оогония через поры оплодотворяющие отроги. При этом один антеридий может оплодотворить одну или несколько яйцеклеток, переливая в каждую некоторое количество содержимого с одним ядром. Имеются гомо-талличные виды, у которых половой процесс происходит между оогониями и антеридиями, образовавшимися на одном и том же мицелии, а есть и гетероталличные. У грибов из родов афаномицес, диктиухус; лептолегния, бреви-легния, геолегния в оогонии образуется только по одной яйцеклетке. Оплодотворенные яйцеклетки во всех случаях одеваются толстыми оболочками и прорастают после периода покоя. [20]
Ооспоры возникают по одной в каждом оогонии. Они имеют разнообразную структуру внешней оболочки - сетчатую, бородавчатую, шиповатую. Ооспора зимует и при благоприятных условиях прорастает с образованием зооспор или толстой ростковой трубки с вздутием на вершине, в котором формируется 40 - 60 зооспор. Но часто везикулы не развиваются, и в этих случаях ооспора прорастает и образует зачаточный мицелий. Таким образом обеспечивается ежегодное возобновление и циркуляция цистопового гриба в природе. [21]
Химанталия интересна тем, что в оогониях развивается по одной очень крупной яйцеклетке ( длиной до 0 5 мм), и тем, что после выхода гамет растение обычно отмирает целиком. В отличие от других фукусовых яйцеклетка химан-талии при развитии в слоевище спорофита уплощается, становится вогнутой с нижней стороны и прикрепляется к грунту краями. Позднее с нижней стороны вырастают нитевидные ризоиды. [22]
Женский репродуктивный орган у красных водорослей - оогоний называют к а р п о г о-ном. Это особая клетка, густо заполненная цитоплазмой и лишенная хлоропласта. Лишь у примитивных представителей флоридеевых и у бангиевых карпогон содержит окрашенный хлоропласт. Нижняя, или брюшная, часть клетки имеет конусовидную форму, верхняя вытягивается, образуя более или менее длинный трубчатый вырост. Вырост служит для улавливания спермациев, называют его т р и-хогиной. Благодаря трихогине карпогон легко отличить от остальных клеток слоевища. Ядро карпогона, как правило, находится в брюшной части. Трихогина может быть короткой и кеглевидной или длинной, узкой и спирально закрученной. У бангиевых собственно трихогина еще отсутствует, а иногда карпогоны имеют лишь короткие сосочкообразные выросты - прообраз будущей трихогины. [23]
У некоторых видов клетки-кормилицы происходят из той же оогонии, из которой образуется соединенный с ними ооцит. Например, у эмбриона Drosophila оогония претерпевает четыре митотических деления, в результате которых образуются 16 клеток. В клетках-кормилицах происходит многократная репликация ДНК без деления самой клетки, поэтому каждая клетка постепенно достигает очень больших размеров, а количество ДНК в ней в тысячу раз превосходит обычную величину ( такая ДНК находится в политенных хромосомах; см. разд. Все 15 клеток-кормилиц, содержащих сотни или тысячи эквивалентов генома, синтезируют вещества, необходимые для одной-единственной яйцеклетки. [24]
Первичные половые клетки мигрируют в формирующуюся гоиаду и превращаются в оогонии; после периода митоти-ческого размножения оогонии дифференцируются в ооцкгы первого порядка, которые приступают к первому делению мейоза. Происходит репликация ДНК, и каждая хромосома состоит после этого из двух хроматид; гомологичные хромосомы конъюгируют по всей своей длине, и между хроматидами спаренных хромосом осуществляется кроссинговер. На этой стадии профаза приостанавливается на то или иное время, которое может составлять от нескольких дней до многих лет в зависимости от вида организма. В этой фазе ооциты первого порядка приобретают наружные оболочки и кортикальные гранулы, накапливают рибосомы, информационную РНК, желток, гликоген и липиды и готовятся к развертыванию программы развития. Во многих ооцитах такая активность отражается на видимой структуре все еще спа-реиных хромосом: они деспирализуются и образуют боковые петли, приобретая характерный вид ламповых щеток, свойственный хромосомам, активно осуществляющим синтез РНК ( см. разд. [26]
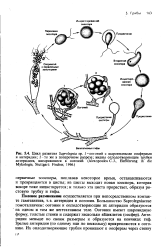
Половое размножение осуществляется при непосредственном контакте гаметангиев, т.е. антеридия и оогония. Большинство Saprolegniaceae гомоталличны: оогонии и оплодотворяющие их антеридии образуются на одном и том же вегетативном теле. Антеридии меньше по своим размерам и образуются на кончиках гиф. Зрелые антеридии ( по одному или по нескольку) прикрепляются к оого-нию. [28]
Однако он своеобразен, так как антеридий, в отличие от оогония, содержащего яйцеклетки, не дифференцирован на гаметы, а просто переливает часть своего содержимого с ядром в яйцеклетку через оплодотворяющий отрог. [29]
В период размножения путем митозов увеличивается число диплоидных половых клеток - оогоний; после прекращения митозов и репликации ДНК в премейотич. В начале периода роста ( фаза медленного роста, или пре-вителлогенеза) ооцит увеличивается незначительно, в его ядре происходят конъюгация гомологичных хромосом и крос-синговер. В цитоплазме увеличивается кол-во органоидов. Эта фаза у ряда животных и человека может длиться годами. В фазе быстрого роста ( вителло-генеза) объем ооцитов увеличивается в сотни, а иногда и тысячи раз, в осн. В период созревания происходят 2 деления мейоза; в результате 1-го деления образуется небольшое полярное тельце и крупный ооцит второго порядка. Мейоз завершается выделением второго полярного тельца и образованием гаплоидного яйца. Полярные тельца впоследствии дегенерируют. Рост ооцитов может проходить по солитарному типу ( без участия спец. Последний объединяет нут-римецтарпый О. [30]
Страницы: 1 2 3 4 5