Зависимость - скорость - окисление
Cтраница 2
Зависимость скорости окисления сульфидов от величины рН исходного раствора приведена в таблице, откуда видно, что гидросульфид и сульфид натрия окисляются очень медленно, причем с переходом от сероводорода к гидросульфиду способность растворов к окислению падает и снова возрастает с переходом от гидросульфида к сульфиду. Основным продуктом окисления при всех испытанных значениях рН исходных растворов является тиосульфат. Сульфит среди продуктов окисления появляется при окислении растворов с рН 9 и выше. [16]
Зависимость скорости окисления сульфидов от величины рН исходного раствора приведена в таблице, откуда видно, что гидросульфид и сульфид натрия окисляются очень медленно, причем с переходом от сероводорода к гидросульфиду способность растворов к окислению падает и снова возрастает с переходом, от гидросульфида к сульфиду. Основным продуктом окисления при всех испытанных значениях рН исходных растворов является тиосульфат. Сульфит среди продуктов окисления появляется при окислении растворов с рН 9 и выше. [17]
Зависимость скорости окисления различных металлов от ориентировки, обнаруженная Тамманом на железе и меди [14, 15], впоследствии неоднократно изучалась. Установлено, что соотношения скоростей окисления различных граней при низких и высоких температурах не одинаковы. По-видимому, это связано с различиями в механизме процесса окисления при изменении температуры. [18]
Выявлена зависимость скорости окисления от размера образцов. [19]
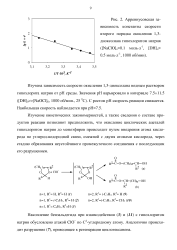 |
Аррениусовская зависимость константы скорости второго порядка окисления 1 3-диоксолана гипохлоритом натрия ( [ NaClO ] моль-л 1, [ DH ] 0 моль-л 1, 1000 об / мин. [20] |
Изучена зависимость скорости окисления 1 3-диоксолана водным раствором гипохлорита натрия от рН среды. Значения рН варьировали в интервале 7.5 - - 11.5 ( [ DH ] 0 [ NaOCl ] o, 1000 об / мин. С ростом рН скорость реакции снижается. [21]
Характер зависимостей скорости окисления кокса от его содержания на катализаторе крекинга, согласно разным литературным источникам, неодинаков. Эти данные в дальнейшем не нашли подтверждения у других исследователей. [22]
Существование зависимости скорости окисления эластомеров от толщины образца позволяет предположить изменение закономерностей окисления при сжатии и растяжении резин, так как эти деформации связаны с увеличением и уменьшением толщины образца. [23]
Факт зависимости скорости топливного окисления от условий сократительного процесса поднимает два вопроса: 1) какова связь между мобилизацией топлива из резервов организма и скоростью использования топлива мышцей и 2) каким образом взаимосвязаны топливные и энергетические потребности мышечной ткани и других тканей. Из числа порождаемых кровью видов топлива только жирные кислоты и глюкоза представляют собой существенные источники энергии для длительной мышечной активности. Тот факт, что уровень глюкозы & крови остается относительно постоянным при изменении скорости работы и потребления топлива, позволяет предположить, что ключевой момент регуляции при энергетическом метаболизме - содержание глюкозы в крови. Основная причина этого обстоятельства состоит, несомненно, в том, что некоторые ткани ( например, мозговая и почечная) в качестве источника топлива опираются только иа глюкозу, так что уровень глюкозы в крови должен оставаться постоянным при различных видах активности. Важнейший процесс, определяющий скорость поступления глюкозы в кровь, - превращение в клетках печени гликогена в глюкозу. В свою очередь уровень Глюкозы в крови связан со скоростью гликолиза и окислительной фосфоризации в мышечной клетке и посредством этого со скоростью использования АТФ и скоростью-мышечного сокращения. Таким образом, благодаря таким связям образуются энергетические метаболические пути между двумя различными тканями - печенью и мышцей. [24]
Факт зависимости скорости топливного окисления от условий сократительного процесса поднимает два вопроса: 1) какова связь между мобилизацией топлива из резервов организма и скоростью использования топлива мышцей и 2) каким образом взаимосвязаны топливные и энергетические потребности мышечной ткани и других тканей. Из числа порождаемых кровью видов топлива только жирные кислоты и глюкоза представляют собой существенные источники энергии для длительной мышечной активности. Тот факт, что уровень глюкозы в крови остается относительно постоянным при изменении скорости работы и потребления топлива, позволяет предположить, что ключевой момент регуляции при энергетическом метаболизме - содержание глюкозы в крови. Основная причина этого обстоятельства состоит, несомненно, в том, что некоторые ткани ( например, мозговая и почечная) в качестве источника топлива опираются только на глюкозу, так что уровень глюкозы в крови должен оставаться постоянным при различных видах активности. Важнейший процесс, определяющий скорость поступления глюкозы в кровь, - превращение в клетках печени гликогена в глюкозу. В свою очередь уровень глюкозы в крови связан со скоростью гликолиза и окислительной фосфоризации в мышечной клетке и посредством этого со скоростью использования АТФ и скоростью мышечного сокращения. Таким образом, благодаря таким связям образуются энергетические метаболические пути между двумя различными тканями - печенью и мышцей. [25]
Что касается зависимости скорости окисления от давления кислорода, то при малых давлениях О2 можно пренебреч. [26]
Была измерена зависимость скорости окисления тетрахлорэтилена от концентрации растворенного кислорода. По закону Генри концентрация растворенного кислорода пропорциональна его парциальному давлению. В качестве инертного разбавителя для кислорода был выбран аргон. Через жидкость при 90 С и мощности дозы 4 72 - 1015 зв / см3 - сек пропускали смесь кислорода и аргона различного состава со скоростью 0 8 л / мин. [27]
Поэтому на зависимость скорости окисления олефина от концентрации гидроперекиси 2-циклогексена влияет присутствие кислоты. [28]
Рассмотрим изучение зависимости скорости окисления 2 3-диметил - 1 4-нафто-хинол - 1 -фосфата ( ДНХФ) перекисью водорода, которое катализируется перо-ксидазой ( HRP), от концентрации ДНХФ. [29]
Рассмотрим изучение зависимости скорости окисления 2 3-дпметнл - 1 4-нафто-хинол - 1 -фосфата ( ДНХФ) перекисью водорода, которое катализируется перо-ксидазой ( HRP), от концентрации ДНХФ. [30]
Страницы: 1 2 3 4 5