Концевые остатки
Cтраница 3
Кроме того, рассматривались частоты встречаемости трех остатков на каждом конце ( а не только одного остатка по модели Зимма - Брэгга), поскольку каждый из трех концевых остатков а-спирали образует только одну водородную связь. Поскольку переходы беспорядочный клубок - - складчатый лист в синтетических полимерах значительно менее изучены, то таких корреляций для - структур пока установить не удалось. [31]
Химические свойства полисахаридов обусловлены особенностями строения их молекулы: большинство остатков моносахаридов израсходовали свои полуацетальные гидроксилы на образование гликозидных связей, а свободные альдегидные группы могут находиться только у концевых остатков. Поэтому восстанавливающие свойства у полисахаридов проявляются в ничтожной степени. [32]
Единственное препятствие, могущее возникнуть в отдельных случаях, состоите том, что некоторая заготовка ( или несколько заготовок), фигурирующая в нанесенных на линейке комбинациях, получится в избытке из одних только концевых остатков, раскраиваемых по линейке при получении остальных заготовок. [33]
Холодная объемная штамповка является одним из наиболее прогрессивных способов изготовления деталей, так как одновременно со снижением трудоемкости дает большую экономию металла, позволяя получать детали практически либо совершенно без отходов, не считая небольших концевых остатков от прутков ( полос) или бунтов, либо с незначительными отходами в виде высечек при пробивке тонких перемычек или дна, а также при доделочных механических операциях. Прочность деталей полученных холодной объемной штамповкой, выше, чем деталей, полученных обработкой резанием, так как волокна материала не перерезаются, а располагаются параллельно поверхности детали, а также выше прочности исходного материала вследствие наклепа, вызванного пластической деформацией. [34]
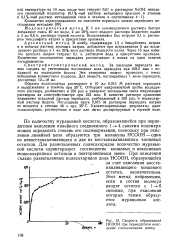 |
Скорость образования НСООН при периодатном окислении глюкоманкана осины. [35] |
По количеству муравьиной кислоты, образовавшейся при перио-датном окислении линейного соединенного 1 - 4 связями полимера можно определить степень его полимеризации, поскольку при окислении линейной цепи образуются три молекулы НСООН - одна из невосстанавливающего и две из восстанавливающего концевых остатков. Для разветвленных полисахаридов количество муравьиной кислоты характеризует соотношение концевых и неконцевых моносахаридных остатков в повторяющемся звене. [36]
Расщепление по Смиту позволяет установить, в каком порядке распределены заместители в молекулах арабоксилана. Концевые остатки арабинозы полностью разрушаются при действии перио-дата, но защищают от окисления замещенные в положении Сз остатки ксилопираноз. При окислении полисахарида с распределением заместителей типа I, после восстановления и мягкого гидролиза образуется с большим выходом ксилопиранозилглицерин. Полисахарид с заместителями типа II в конечном итоге должен дать ксилопиранозилглицерин и ксилобиозилглицерин, а из полисахарида типа III должны образоваться гликозиды глицерина и олиго-сахаридов, размер которых определяется числом звеньев с рядом расположенных заместителей. [37]
Концевые остатки ткани ( табл. IV-46) образуются вследствие некратности длин кусков ткани длинам выкраиваемых деталей, а также из-за вырезки дефектов ткани, не допускаемых в изделиях мебели. Нормативы отходов концевых остатков учитываются только при выделении фондов на ткань и в нормы расхода на изделие не включаются. [38]
 |
Спиралевидное строение макромолекулы амилозы. штриховка - соединения включения. а - молекула иода / я. [39] |
В крахмальном зерне амилоза располагается, главным образом, внутри, а оболочка состоит из амилопектина. Некоторая часть концевых остатков глюкозы этерифицированы фосфорной кислотой. [40]
Для определения С-концевых остатков чаще всего используют ферментативный гидролиз карбокснпептидазами, к-рые специфически расщепляют пептидные связи, образованные С-коицевыми остатками. Поскольку после отщепления концевых остатков фермент атакует послед, пептидные связи, измерение скорости отщепления отдельных аминокислот позволяет анализировать также и С-концевую аминокислотную последовательность. [41]
В течение последних десятилетий было показано, что белки являются природными полимерами, состоящими из длинных полимерных цепей, повторяющимися звеньями в которых служат а-аминокислотные остатки, поэтому изучение инфракрасных спектров аминокислот имеет очень большое значение для понимания структуры белка и его функции. Особенно большое значение имеет определение концевых остатков и значение чередования различных остатков аминокислот в полимерной цепи. [42]
Далее, термин неупорядоченная, или статистическая, атака, строго говоря, неправомочен и скорее относится к атаке на срединные связи, удаленные от концов полимерного субстрата. Это вызвано тем, что константа ассоциации концевых остатков субстрата с активным центром ( точнее, лишь с частью активного центра) должна быть ниже, чем со всей сорбционной областью, и соответственно вероятность атаки потенциально расщепляемой связи у концов полимерного субстрата ниже, чем вероятность атаки любой связи в середине полимера. В этом смысле атака протона по отношению к олигомерному субстрату должна быть значительно более неупорядоченной, чем атака фермента. [43]
Обычно протеазы разделяют на пептидазы и протеи-назы. Первые катализируют расщепление тех пептидных связей, которые соединяют концевые остатки аминокислот с главной полипептидной цепью, а вторые - пептидные связи, удаленные от концевых остатков аминокислот в пептидной цепи. [44]
Ветвление повышает растворимость гликогена. Кроме того, благодаря ветвлению создается большое количество невосстанавливающих концевых остатков, которые являются местами действия гликогенфосфорилазы и гликогенсинтазы. [45]
Страницы: 1 2 3 4