Гистидиновые остатки
Cтраница 1
 |
Таутомерия пнразолоа н нмидазолов.| Преимущественное образование 4-ннтроструктуры для Ц5 - ннтроими-лазола. [1] |
Гистидиновые остатки обнаружены в активных положениях ри-бонуклеазы и некоторых других ферментов. Действие некоторых ферментов, очевидно, заключается в катализе протонного переноса ( см. также разд. Система имидазола проявляет как кислые, так и основные свойства. [2]
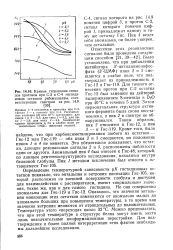 |
Кривые титрования сигналов протонов при С-2 и С-4 гистидиновых остатков рибонуклеазы, соответствующие спектрам на. [3] |
Определение температурной зависимости рК гистидиновых остатков показало, что энтальпия и энтропия ионизации Гис-105, который расположен на внешней поверхности глобулы доступен для взаимодействия с растворителем, имеют нормальные значения, такие же, как и для самого гистидина. [4]
Имеются основания считать, что гем связан с глобиновым компонентом через гистидиновые остатки в полипептидных цепях молекулы гемоглобина. [5]
У этой реакции, сопровождающейся запасанием энергии, есть промежуточный этап, во время которого происходит фосфорили-рование самой молекулы фермента по одному из гистидиновых остатков в его активном центре. Именно богатая энергией фосфатная группа, участвующая в этом фосфорилировании, и переносится на GDP с образованием GTP. Сопряженное образование GTP за счет энергии, выделяющейся при окислительном фосфорилировании а-кетоглутарата, представляет собой еще один пример фосфо-рилирования на уровне субстрата. Вспомним, что нам уже знаком один пример фосфорилирования этого типа, а именно сопряженный синтез АТР за счет энергии, выделяющейся при окислении глицеральдегид-3 - фосфата в ходе гликолиза ( разд. [6]

Приведенные данные свидетельствуют о том, что кроме гидрофобных взаимодействий при образовании комплекса бенздиазепи-ны - ЧСА принимают участие и отрицательные электростатические силы. Следовательно, гистидиновые остатки альбумина не принимают участия при взаимодействии макромолекулы с бенздиа-зепинами, так как при значении рН выше 6 6 они депротонированы, а количество связанных анионных групп транквилизаторов остается неизменным. [8]
Точно так же при денатурации увеличивается число связываемых динитрофенолом гистидиновых групп, равно как и число обнаруживаемых р - и укарбоксильных групп. Так, после денатурации р-лактоглобулина количество реактивных гистидиновых остатков возрастает с двух до четырех, в случае карбоксигемо-глобина освобождаются 36 карбоксильных групп на одну молекулу белка. Увеличивается и интенсивность цветных реакций на тирозин с фосфорномолибденовой кислотой или диазореагента-ми и на аргинин при обработке реактивом Сакагучи. [9]
После инкубации электрофореграмму высушивают и снова подвергают электрофорезу при рН 5 в перпендикулярном направлении. В парах аммиака происходит отщепление карбэтоксильных групп, гистидиновые остатки становятся доступными протонированию, и при электрофорезе пептиды, содержащие гистидин, смещаются от занимаемого другими пептидами диагонального положения в сторону катода. [10]
Бычья панкреатическая рибонуклеаза А была тщательно исследована с помощью ПМР. Однако удалось достоверно идентифицировать только атомы водорода при С-2 имидазола четырех гистидиновых остатков. [11]
Гистоны, как и другие белки, синтезируются в цитоплазме на рибосомах и в процессе миграции в ядро, посадки на ДНК и даже уже в составе хроматина подвергаются многообразным химическим модификациям. В гистонах происходит ацетилирование лизиновых остатков, снимающее их положительный заряд, метилирование лизиновых, аргининовых и гистидиновых остатков, фосфорили-рование треонинов и серинов. К существенному изменению свойств гистонов приводит их поли - АВР-рибозилирование. Иногда к молекулам гистонов в результате модификации присоединяются и более объемистые группы. [12]
Гистоны, как и другие белки, синтезируются в цитоплазме на рибосомах и в процессе миграции в ядро, посадки на ДНК и даже уже в составе хроматина подвергаются многообразным химическим модификациям. В гистонах происходит ацетилирование лизиновых остатков, снимающее их положительный заряд, метилирование лизиновых, аргининовых и гистидиновых остатков, фосфорили-рование треонинов и серинов. К существенному изменению свойств гистонов приводит их поли - АОР-рибозилирование. Иногда к молекулам гистонов в результате модификации присоединяются и более объемистые группы. [13]
Низкоспиновый комплекс образуется лишь в том случае, когда в пятом и шестом положениях находятся группы, расположенные справа в спектрохимическом ряду. Так, в низкоспиновом ферроцитохроме с пятое и шестое координационные положения занимает азот имидазольных групп гистидиновых остатков. При физиологических условиях ферро-цитохром с не может связывать кислород, окись углерода, фтор или циа-нид. Окисленный ферроцитохром с также является низкоспиновым. Миоглобин и гемоглобин представляют собой высокоспиновые комплексы ферро-иона с имидазольной группой гистидина в пятом положении и молекулой воды в шестом. Полный магнитный момент молекулы оксигемоглобина становится равным нулю. Комплекс ферро-иона со спаренными спинами дает темно-красную окраску благодаря поглощению, связанному в основном с порфириновым кольцом. Окисление этих двух белков кислородом протекает сравнительно медленно, несмотря на то, что термодинамически это выгодно. Частично это связано с тем, что каждый такой процесс происходит с изменением спинового числа. При использовании других окисляющих агентов получаются высокоспиновые комплексы ферри-иона с белками, называемые метмиоглобином или метгемоглобином. [14]
По-видимому, отсутствие эффекта кооперативности при модификации гистидина должно отражать постепенную, но при этом ускоряющуюся потерю железо-связывающей активности в зависимости от числа измененных ги-стидинов. Возможно, также, что потеря способности связывать железо отражает изменения в третичной структуре белка при увеличении числа модифицированных гистидиновых остатков. [15]
Страницы: 1 2