Остов - молекула
Cтраница 3
 |
Плотности я-л - и я - я - электрошшх переходов в формальдегиде. [31] |
Наибольшую вероятность имеют такие электронные переходы, при которых геометрия ядерного остова молекулы в исходном и конечном состояниях одинакова. В такой форме принцип Франка - Кондона устанавливает изменение колебательного квантового числа при электронном переходе и тем самым колебательную структуру полосы поглощения. [32]
В проведенном выше рассмотрении предполагалось, что для связей, создающих остовы молекул бензола и бутадиена, вполне достаточно развитых ранее идей о перекрывании атомных орбит, но для я-электронов эти представления непригодны, и тогда возникает вопрос о том, законно ли рассматривать разные электроны в одной и той же молекуле с помощью различных концепций. [33]
Для обсуждения переходов между орбитами, сильно делокализо-ванными по всему остову молекулы, может быть использована теория поля лигандов, применимая таким образом к спектрам переноса заряда и сходным с ними типам спектров. [34]
Это означает, что ридбергов электрон очень слабо связан с ядерным остовом молекулы, который поэтому способен вращаться, не увлекая за собой электрон; см. случай связи Гунда ( d) в разд. Рид-берговы электроны характеризуются малым квантовым дефектом: они настолько диффузны, что почти не взаимодействуют с внутренними электронами, и их волновые функции похожи на волновые функции атома водорода. [35]
О У / С) инвариантны отнсснтельно преобразований точечной группы симметрии ядерного остова молекулы или кристалла. [36]
Реакционная серия должна иметь один и тот же скелет и состав остова молекулы R, включая последовательность простых и кратных связей, если они имеются. [37]
В анализируемую электронную систему не включают а-элек-троны, относя их к остову молекулы. [38]
Было показано, что с увеличением расстояния между асимметрическим атомом и остовом молекулы при постоянной длине нормальной цепи термическая стабильность смектической и холестерической мезофаз зигзагообразно возрастает. При увеличении длины цепи н-алкильного радикала область существования смектической мезофазы расширяется за счет сужения холестерической. Такая зависимость является общим правилом для гомологических серий мезоморфных соединений. [39]
Особенно отчетливы закономерности этого типа в фотохимических реакциях, в которых сохраняется остов молекулы, и обычно речь идет лишь об электронных переходах в возбужденные состояния. [40]
Возможность раздельного учета влияния на колебательные спектры деформации электронной оболочки и изменения строения остова молекулы представляется в некоторой мере проблематичной. Однако можно предположить, что деформация электронной оболочки при сопряжении кратных связей приводит главным образом к изменению полосы электронного поглощения и действие этого фактора на спектры комбинационного рассеяния осуществляется через показатель поглощения. Тогда, пользуясь результатами § 5, можно исключить влияние данного фактора и выделить в спектрах комбинационного рассеяния те изменения, которые непосредственно связаны с изменением строения остова молекулы. Тот факт, что степень деполяризации лишь незначительно меняется при изменении частоты возбуждающей линии ( см. табл. 24), по-видимому, подтверждает приведенные рассуждения. Исключив эти множители, мы можем найти след и анизотропию тензора а, которые не зависят непосредственно от частоты возбуждающего света. [41]
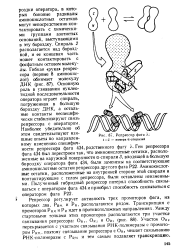 |
Репрессор фага /.. 1 - 5 - номера а-спнралей. [42] |
Спираль 2 располагается над бороздкой, и ее концевая часть может контактировать с фосфатным остовом молекулы. Основную роль в узнавании нуклео-тидной последовательности оператора играет спираль, погруженная в большую бороздку ДНК, а остальные контакты неспецифически стабилизируют связь репрессора с оператором. Аминокислотные остатки, расположенные на внутренней стороне этой спирали и контактирующие с телом репрессора, были оставлены неизменными. [43]
Нуклеиновые кислоты заряжены отрицательно, что обусловлено диссоциацией фосфатных групп, которые в структуре остова молекулы чередуются с молекулами пентозы. [44]
Во второй половине XIX века были разработаны многочисленные методы взашдопревращения функциональных групп и построения углеродного и гетероатомного остова молекул. [45]
Страницы: 1 2 3 4