Первичный ответ
Cтраница 1

Первичный ответ может возникнуть недостаточно быстро, чтобы подавить инфекцию, но уж коль скоро человек заболел и выжил, то в будущем встреча с этим патогеном не будет представлять для него опасности благодаря быстрому вторичному ответу. [2]
Первичный ответ свидетельствует о поступлении сигнала от данного рецептора в корковое представительство анализатора. [3]
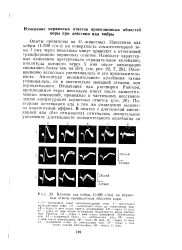 |
Влияние яда кобры ( г / мл на первичные ответы проекционных областей коры. [4] |
А - первичный ответ до аппликации яда, IS - через 5 мин после снятия аппликации, 1В - через 20 мин, I / - через 10 мин после отмывания поверхности коры раствором Рингера, Д - через 5 мин после повторной аппликации лда; 2А - первичный отост до аппликации яда, 2Б - через 7 млн после аппликации, 2В - через 20 мин, 2f - через 30 мшг. ЗЛ - - псрничнып ответ до аппликации яда, iB - через а мин после снятия аппликации, 3 -через 15 мин. [5]
Увеличение амплитуды первичных ответов происходит и при аппликации 1 % раствора атропина. В то же время ранние компоненты ассоциативных ответов, отводимые от передней теменной области, демонстрируют устойчивость к примененным фармакологическим агентам, так же как и к яду кобры. [6]
Напротив, 0 1 % раствор стрихнина вызывает увеличение амплитуды первичных ответов. [7]
Сходная картина наблюдается при применении 0 5 % раствора нембутала, который также обусловливает исчезновение и инверсию негативного отклонения первичных ответов и возрастание длительности и амплитуды положительной волны. [8]
Это свидетельствует о том, что М - иммуноглобулиннесущие В-клет-ки вовлекаются в ответ в том же количестве, что и при первичном ответе. В то же время, более специфичный IgG-ответ развивается интенсивнее и по количеству антител в несколько раз превышает первичный ответ. [9]
Было показано, что в проекционных отделах наложение 1 % раствора ГАМК уже через 30 - 60 сек приводит к типичным преобразованиям первичных ответов. Эти изменения проявляются в исчезновении и инверсии отрицательной волны, а также в соответствующем увеличении длительности протекания и возрастании амплитуды положительной волны. При этом вслед за трансформированными первичными ответами возникает вторичное отрицательное колебание. [10]
По нашим данным, предварительное ( за 2 - 3 ч) введение кошкам больших доз гепарина ( 50 мг / кг) предотвращало угнетение первичных ответов коры при прямой аппликации яда кобры ( 1 %) на поверхность соматосенсорной зоны. Механизм защиты гепарином именно деполяризующих синапсов пока не ясен. Я - МихельсО Н, Э. В. Зеймаль, 1970), что может обусловить относительную устойчивость холинорецептивной мембраны к действию яда. Очевидно, в условиях защиты гепарином деполяризующих синапсов яд в меньшей степени угнетает возбуждающий постсинаптический потенциал апикальных дендритов, что проявляется в устойчивости отрицательного колебания первичных ответов к действию яда кобры. Другой возможный механизм защитного действия гепарина может быть связан с его хорошо выраженными комплексообразующими свойствами ( А. А. Маркосян, 1969), в результате чего инактивация яда происходит путем связывания его гепарином. [11]
Установленный факт устойчивости ранних компонентов ассоциативных ответов к яду кобры был подвергнут специальному анализу с использованием ряда фармакологических агентов, с хорошо изученным возбуждающим или угнетающим действием на первичные ответы. С этой целью были использованы ГАМ. [12]
При 1 ] / 2 - месячной непрерывной затравке кроликов озоном в концентрации 0 05 мг / м3 Н. Д. Экслер совместно с нами, начиная с 4 - й нед опыта, наблюдала у животных сдвиги в картине вызванных потенциалов: снижение амплитуды первичного ответа и медленной отрицательной волны с одновременным удлинением ее. В этом случае следует, по-видимому, говорить о нарушении в системе коркового торможения. Можно полагать, что отмеченная депрессия амплитуды первичного ответа и изменение амплитудно-временных параметров вторичных реакций вызванного потенциала отражают препатологическую реакцию мозга. [13]
Найдено, что в начале затравки происходит усиление процесса коркового торможения, которое можно рассматривать как механизм компенсации, направленной на устранение результатов воздействия токсичного вещества. Обнаруженные в дальнейшем падение амплитуды первичного ответа, медленной отрицательной волны и позднего ответа, а также уменьшение продолжительности медленной отрицательной волны свидетельствуют об ослаблении коркового торможения. Прогностически такой сдвиг можно считать неблагоприятным. То обстоятельство, что изменение вызванных потенциалов развивалось параллельно в гиппокампе и зрительной коре, указывает, по-видимому, на общий механизм действия токсичных веществ в низких концентрациях при длительном их поступлении через кровь на синаптиче-ские структуры различных образований головного мозга. [14]
В рецепторах катехоламина, например, первичный ответ состоит в продуцировании вторичного мессенджера сАМР, который с помощью протеинкиназы регулирует не только ионную проницаемость возбудимых мембран, но также энергию метаболизма и биосинтез белка в клетке. Рецепторы, определяемые как молекулы, связывающие эндогенные лиганды, являются в действительности компонентами мембранных комплексов, состоящих из молекул разных видов: одни из них связывают лиганды, а другие функционально активны в мембране. Способ, с помощью которого регулируется ионная проницаемость клеточной мембраны, можно рассмотреть на примере модели, разработанной для аксональных ионных каналов ( гл. [15]
Страницы: 1 2