Большинство - водоросль
Cтраница 2
В фотосинтезирующих клетках активные пигменты расположены внутри ламеллярных мембран в виде функционально-организованных единиц. У фотосинтезирующих эукариот ( высших растений и большинства водорослей) несущие пигмент мембраны заключены в специфических органеллах - хлоропла-стах. У высших растений морфологические различия между хлоропластами невелики, в то время как у водорослей форма и размеры хлоропластов значительно варьируют. Chlorella, например, имеет единственный чашевидный хлоропласт, тогда как хлоропласты некоторых видов Spirogyra представляют собой длинные, спирально закрученные образования, лежащие вдоль всей клетки. [16]
Несмотря на возросший потенциал развития биологических обрастаний в тешюобменной аппаратуре при использовании городских сточных вод, можно получить хорошие результаты по предотвращению этих биообрастаний, применяя биоцидную обработку воды. При применении хроматных ингибиторов замедляется развитие грибов, большинства водорослей ( в градирнях) и бактерий ( в теплообменных аппаратах), поскольку соли хрома являются токсичными. Особенно эффективно соли хрома подавляют развитие сульфатовосстанавливающих бактерий в теплообменных аппаратах. Фосфатные ингибиторы являются благодатной средой для развития микроорганизмов. Если городские сточные воды после полной биологической очистки используются в качестве добавочных вод без биоцидной обработки, может происходить интенсивный процесс коррозии даже при применении хроматных или нехроматных ингибиторов. [17]
Для водорослей как фототрофных организмов первым условием существования является наличие света, источников углерода и минеральных веществ, а основной средой жизни для них служит свободная вода. Однако это справедливо не для всех, а только для большинства водорослей и не исчерпывает всего многообразия наблюдающихся у них способов питания и мест поселения. [18]
Вопрос о происхождении и эволюции водорослей очень сложен из-за разнообразия этих растений, особенно их субмикроскопического строения и биохимических признаков. Решение этой проблемы затруднено, кроме того, плохой сохранностью большинства водорослей в ископаемом состоянии и отсутствием связующих звеньев между современными отделами растений в виде организмов промежуточного строения. [19]
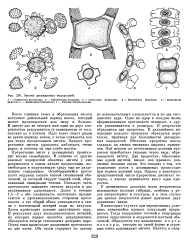 |
Зиготы десмидиевых водорослей. [20] |
Одно из ядер в каждом вновь сформированном протопласте отмирает, а другое увеличивается в размерах. В результате образуются два проростка. В дальнейшем посередине каждого проростка образуется перетяжка, типичная для большинства десмидиевых водорослей. Оболочка проростка всегда гладкая; он лишен типичных видовых признаков. При первом же вегетативном делении проросток приобретает типовые черты вида, образовавшего зиготу. Два проростка, образованные одной зиготой, имеют, как правило, различные половые знаки. Это говорит о том, что разделение полов у десмидиевых происходит при первом делении ядра. Однако у некоторых представителей число проростков достигает четырех, а иногда развивается всего один проросток. [21]
Современными исследоглкиями установлено, что вполне сфор-ифованный хлоропласт иг с - т двойную липопротеидную меморану. Внутри хлоропласта находится бес - ветная белковая строма ( матрикс), в которую погружены содержа - - щие пигменты ламеллярные системы, различные у разттт растений. Так, у высших растений они образуют граны; у большинства водорослей грчн нет, но ламеллы проходят через весь хлоропласт пронизывают его. Кроме ламеллярных систем в матриксе хлоропласта находятся другие структурные элементы: осмиофильные глобулы, рибосомы. [22]
Первичные фотосинтезирующие жгутиковые эволюционировали по двум главным направлениям. Второе направление эволюции жгутиковых характеризуется преобладанием зеленой окраски и наличием хлорофилла Ъ наряду с хлорофиллом а. Хлоропласты желто-зеленых водорослей не содержат хлорофилла Ъ и имеют такое же строение, как у большинства водорослей с бурыми пигментами. В качестве запасного вещества может образовываться хри-золаминарин; крахмал и парамилон отсутствуют. В пределах каждого из этих направлений эволюция пошла по нескольким линиям, нередко параллельно, сопровождаясь возникновением сходных форм строения тела. [23]
Активная реакция среды измеряется с помощью лабораторного рН - метра типа ЛПУ. Обычно такие приборы позволяют определять рН растворов с точностью до десятых долей. В качестве электродов употребляются стеклянные и вспомогательные ( хлоркалиевые) электроды. Стеклянный электрод должен предварительно вымачиваться в дистиллированной воде не менее суток. Во вспомогательный электрод предварительно заливается насыщенный раствор хлористого калия. Растворы, в которых надо измерить рН, заливаются в специальные стаканчики, которые подставляются под электроды и после установки стрелки гальванометра проводится измерение. Такие измерения рН необходимо производить через определенные промежутки времени в течение всего опыта. Подщелачивание среды, происходящее в процессе естественного роста большинства водорослей, указывает на нормальное развитие их и часто сопровождается увеличением их численности. [24]
Страницы: 1 2