Плазмида
Cтраница 2

Размеры плазмид варьируют от менее 1 до более 500 т.п.н. Каждая из них содержит сайт начала репликации ( ori), без которого репликация плазмиды в клетке-хозяине была бы невозможна. [17]
 |
Токсичность природных и рекомби-нантных подвидов В. thuringiensis для насекомых Pieris brassicae ( капустница, Aedes aegypti ( комар и Phaedon cochleariae ( жук 1. [18] |
Нестабильность плазмид еще раз убеждает в целесообразности интеграции клонированного cry - гена ( генов) с хромосомной ДНК. Рассмотрим один успешный пример такой интеграции. [19]
Строение плазмиды pBR322 показано на рис. 7.3. Она несет гены устойчивости к тетрациклину и ампициллину. [20]
Репликация плазмид происходит независимо от репликации хромосомы, причем на одну хромосому в бактерии может приходиться одна или несколько одинаковых плазмид. Эписомы - это плазмиды, способные включаться в хромосому бактерии. Некоторые внехромосомные элементы могут вести себя как эписомы в одних бактериях и как плазмиды в других. Плазмиды могут быть инфекционными ( переносимыми) или неинфекционными. В первом случае они содержат гены для синтеза - половых пилей ( разд. А, 1 г) и способны - переносить свою ДНК в другую клетку. Если плазмида способна интегрироваться с хромосомой, а потом и выходить из нее, захватывая с собой при этом другие гены, то такую плазмиду называют фактором пола. Выше уже шла речь о том, что процесс переноса генов с помощью фактора F был широко использован при картировании бактериальных хромосом. [21]
Размеры плазмид и эписом могут быть различными. Половой фактор F-1 представляет собой кольцевую суперспирализованную ДНК, мол. Некоторые более крупные колициногенные факторы одновременно являются и половыми факторами. Плазмида переносит также ген ( или гены), придающий устойчивость по отношению к токсинам, в бактериальную клетку-хозяина. [22]
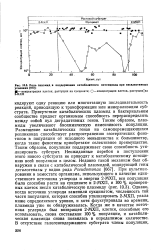 |
Роль плазиид в поддержании катаболитного потенциала при неселективных условиях. [23] |
Присутствие катаболических плазмид в бактериальном сообществе придает организмам способность перераспределять между собой пул деградативных генов. Таким образом, плаз-миды увеличивают биохимическую изменчивость популяции. Размещение катаболических генов на самоперемещающихся репликонах способствует распространению эзотерических фенотипов в популяции от исходного меньшинства к большинству, когда такие фенотипы приобретают селективное преимущество. Таким образом, популяция поддерживает свою способность утилизировать субстрат. Неожиданные перебои в поступлении этого нового субстрата не приводят к метаболическому истощению всей популяции в целом. Однако, когда источник углерода заменили сукцинатом, численность той части популяции, которая способна утилизировать 2 - МХП, упала ниже определяемого уровня, и хотя флуктуировала во времени, плазмида уже не обнаруживалась. Когда же в качестве источника углерода был возвращен 2 - МХП, клетки, способные его использовать, снова составляли 100 % популяции, и катаболи-ческая плазмида снова появилась в определимом количестве. [24]
Пластичность катаболических плазмид обеспечивает механизм, с помощью которого обмен генетическим материалом может привести к созданию организма, способного к эффективной утилизации нового субстрата в фазе обогащения, как это описано для организмов, использующих галогензамещенные жирные кислоты. На этом основании рассматриваются две возможности, которые могли возникнуть в процессе обмена генетическим материалом между двумя организмами в исходной обогащенной культуре. С одной стороны, могло произойти накопление дегалогеназных структурных генов в исходной плаз-миде, которая претерпевает дальнейшую структурную реорганизацию, перемещаясь от штамма к штамму. И наоборот, возможно также, что эти гены оказались связаны с пятью различными плазмидами. [25]
В плазмиду можно встроить участок ДНК, взятый откуда угодно, скажем, ген человека, и она внутри бактерии начинает вырабатывать белок, соответствующий человеческому гену. Это и есть тот трюк, который генные инженеры научились проделывать с проворством искусных магов. При этом используется один из трех приемов. [26]
В экспрессирующую плазмиду встроен ген, в котором сайт узнавания эндонуклеазы рестрикции EcoRl начинается с G, входящего в триплет первого метионина. Встройте в этот белок дополнительный трипептид. Какие ограничения налагаются на строение этого трипептида. [27]

Этой плазмидой трансформируют культуру дефектных по тимидинкиназе животных клеток, обычно фибробластов куриного эмбриона, предварительно инфицированных ВКО дикого типа, который синтезирует функциональную тимидинкиназу. [29]
О плазмидах заговорили, причем не столько молекулярные биологи, сколько медики, после того как в 1959 г. японские исследователи обнаружили, что неэффективность хорошо зарекомендовавших себя антибиотиков при лечении дизентерии у некоторых больных обусловлена тем, что бактерии, которыми заражены эти пациенты, несут в себе плазмиду, содержащую сразу несколько генов устойчивости к разным антибиотикам. [30]
Страницы: 1 2 3 4 5