Плазмида
Cтраница 3
Сконструированная экспрессионнеая плазмида была введена в геном микроорганизма-хозяина с помощью стандартной процедуры трансформации. [31]
Выяснено существование плазмид, ингибирующих F-фактор, а также несовместимых с другими, введенными в одну и ту же клетку. Доказано удвоение числа плазмид репликацией, а также возможность присутствия в одной бактериальной клетке не только двух, но и большего числа плазмид. При исследовании полового фактора были получены плазмидные мутанты, известные теперь также и у R-факторов. По-видимому, мутации возможны у всех плазмид. [32]
При выделении плазмид для увеличения выхода плаз-мидной ДНК к культуре за несколько часов до сбора клеток иногда добавляют хлорамфеникол. [33]
Значительная изменчивость ката-болических плазмид в этом роде объясняет широкие катаболические возможности, которыми обладают его представители. Физический размер этих плазмид позволяет им кодировать большое количество генов. ДНК в количестве, достаточном для кодирования приблизительно 150 генов. Это означает, что даже в случае, когда с данной плазмидой сцеплен ряд фенотипических маркеров, таких как множественная устойчивость к лекарственным препаратам [668] или 12 катаболических ферментов, которые катализируют расщепление толуола и м - и n - ксилолов ( плазмида TOL [648]), большие сайты плазмиды кодируют неидентифицированные фенотипические признаки. Следовательно, наши знания даже об относительно хорошо изученной архетипической TOL-плазмиде ( pWWO [669, 670]), ограничены областью ката-болической функции, репликации и переноса плазмиды. [34]
После этого линейную плазмиду, обладающую хвостами, и кДНК просто смешивают и предоставляют им возможность образовать пары между комплементарными основаниями. Среди продуктов такой гибридизации будет находиться увеличенная кольцевая плазми-да, содержащая новый ген, который связан с плазмидой только за счет комплементарного взаимодействия липких концов. Эти концы можно теперь ковалент-но соединить с помощью ДНК-лигазы ( разд. [35]
 |
Использование терминальной транс-феразы для достраивания концов ДНК с целью обеспечения фрагментов ДНК комплементарными липкими концами. [36] |
В каждой плазмиде содержится от 2000 до 100000 оснований. Маленькие плазмиды могут присутствовать в клетке в количестве 20 и более копий; плазмид более крупного размера в клетке бывает не более 1 - 2 копий. Каждая плазмида содержит несколько, а иногда и много генов, которые реплицируются, транскрибируются и транслируются независимо от хромосомных генов, но одновременно с ними. Плазмиды легко выделить и отделить от бактериальных хромосом, от которых они отличаются по размеру, нуклеотидному составу и плотности. Плазмиды обладают двумя замечательными свойствами, полезными для генетического манипулирования. Во-первых, они могут переноситься от одной клетки к другой и даже от бактерии одного вида к бактерии другого. [37]
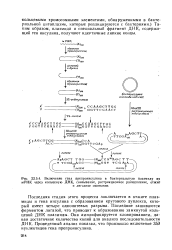 |
Включение гена препроинсулина в бактериальную плазмиду из мРНК через кольцевую ДНК, связывание, рестрикционное расщепление, отжиг. [38] |
Таким образом, плазмида и специальный фрагмент ДНК, содержащий ген инсулина, получают идентичные липкие концы. [39]
Для своей репликации плазмиды используют репликативную машину клетки-хозяина, однако репликация плазмид происходит независимо от хромосомы. Каждая плазмида является самостоятельным репликоном, сама контролирует собственную репликацию и поддерживается в клетке в определенном, характерном для нее числе копий. Для характеристики плазмидных репликонов их принято разбивать на группы несовместимости. [40]
Однако стратегия выживания плазмид, как и в случае транспозонов, основывается не только на том, чтобы приносить пользу клеткам-хозяевам. Прежде всего сохранению плазмид в популяциях бактерий способствует высокая стабильность их наследования. Она достигается точным контролем репликации и секрегации плазмид по дочерним клеткам ( см. гл. II), но многие плазмиды имеют еще дополнительные приспособления для повышения стабильности наследования. Другая плазмида, R1 ( кодирующая устойчивость к нескольким антибиотикам), достигает той же цели другим путем. Один из генов этой плазмиды кодирует токсичное для клетки вещество, а другой ген - короткоживущий ингибитор этого вещества, так что наличие плазмиды для клетки не вредно. [41]

Чтобы предотвратить утрату плазмиды, была предусмотрена интеграция участка AOXlp - HBsAg-AOXlt в геном P. [43]
Однако стратегия выживания плазмид, как и в случае транспозонов, основывается не только на том, чтобы приносить пользу клеткам-хозяевам - Прежде всего сохранению плазмид в популяциях бактерий способствует высокая стабильность их наследования. Она достигается точным контролем репликации и секрегации плазмид по дочерним клеткам ( см. гл. II), но многие плазмиды имеют еще дополнительные приспособления для повышения стабильности наследования. Другая плазмида, R1 ( кодирующая устойчивость к нескольким антибиотикам), достигает той же цели другим путем. Один из генов этой плазмиды кодирует токсичное для клетки вещество, а другой ген - короткоживущий ингибитор этого вещества, так что наличие плазмиды для клетки не вредно. [44]
Обычно о присутствии плазмид в бактериальной клетке судят по проявлению определенных признаков, к которым относится устойчивость к отдельным лекарственным препаратам, способность к переносу генов при конъюгации, синтез веществ антибиотической природы, способность использовать некоторые сахара или обеспечивать деградацию ряда веществ. Из перечисленного выше видно, что плазмиды делают возможным существование организмов в более широком диапазоне условий внешней среды, т.е. действуют как факторы адаптации. Большую группу составляют плазмиды с нерасшифрованными функциями; такие плазмиды выявляют с использованием физико-химических методов. [45]
Страницы: 1 2 3 4 5