Задержка - деление
Cтраница 2
Приблизительно такой размер имеет ядрышко. Если причина задержки деления заключается в нарушении цикла нуклеиноной кислоты, то не исключена возможность, что ядрышко имеет к этому отношение, так как на стадии покоя оно служит резервуаром рибонуклеиновой кислоты. [16]
Задержка деления, а не его полное подавление, означает, что в этом случае мы встречаемся с таким действием радиации, от которого клетка оправляется. Поэтому всякое объяснение результатов опытов по задержке делений должно предусматривать какой-то механизм восстановления. [17]
У многих объектов облучение вызывает задержку деления, и часть клеток, приступающих к делению после этой задержки, гибнет. Этот факт наводит на мысль, что задержка деления и гибель клеток при делении, возможно, представляют собой по существу сходные явления, различающиеся скорее количественно, чем качественно. Согласно этому предположению умеренное повреждение клетки вызывает некоторую задержку деления или затягивание его ранних стадий, тогда как более сильное повреждение, вызванное облучением в более сильных дозах, ведет к большей задержке деления и к гибели клетки при делении. Такое толкование не является, однако, обязательным и, по-видимому, в ряде случаев задержка деления и гибель клеток при возобновлении деления представляют собой независимые явления. Общий метод проверки независимости или связанности этих явлений заключается в том или ином изменении условий опыта, например типа излучения, его интенсивности, температуры, с тем, чтобы установить, носят изменения этих двух эффектов сходный или различный характер. [18]
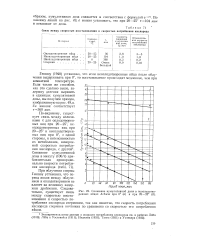 |
Связь между скоростью восстановления и скоростью потребления кислорода.| Снижение кумулятивной дозы в неоплодотворенных яйцах Arbacia при 0 ( а и при 20 - 25 ( б. [19] |
Геншоу ( 1940) установил, что если неоплодотворенные яйца после облучения выдерживать при 0, то восстановление происходит медленнее, чем при комнатной температуре. Если таким же способом, как это сделано выше, задержку деления выразить в единицах кумулятивной дозы, мы получим прямую, изображенную на рис. 49 а. Ее наклон соответствует т360 мин. [20]
VI, если в клетках процесс деления к моменту облучения уже настолько продвинулся, что ( в отличие от клеток на более ранних стадиях) они не испытывают задержки деления, то облучение может привести к появлению в метафазе и анафазе картин, указывающих на клейкость поверхности хромосом. При этом в метафазе хромосомы слипаются, а разъединение сестринских хроматид в анафазе оказывается затрудненным. [21]
Для наших целей ее можно определить как дозу облучения в рентгенах, которая, если бы она была сообщена клетке, мгновенно привела бы ее в то же состояние ( в отношении химического изменения, ответственного за задержку деления), в котором клетка находится в данный момент. Представление о кумулятивной дозе использовали многие исследователи ( Гофман и Рейнгард, 1934; Куимби и Маккомб, 1937; Ли, 1938а, Ь) при разработке теорий тех действий излучения, которые допускают восстановление и в отношении которых ответственные за эффект химические изменения пока неизвестны. Если окажется, что химическое изменение заключается в выработке под влиянием облучения какого-то вещества, подавляющего деление, то кумулятивная доза будет пропорциональна концентрации этого вещества. Если же ответственное за эффект изменение заключается в разрушении какого-то компонента клетки, а возврат к норме - в его восстановлении, то кумулятивная доза будет пропорциональна недостатку этого компонента в клетке в данный момент. [22]
У многих объектов облучение вызывает задержку деления, и часть клеток, приступающих к делению после этой задержки, гибнет. Этот факт наводит на мысль, что задержка деления и гибель клеток при делении, возможно, представляют собой по существу сходные явления, различающиеся скорее количественно, чем качественно. Согласно этому предположению умеренное повреждение клетки вызывает некоторую задержку деления или затягивание его ранних стадий, тогда как более сильное повреждение, вызванное облучением в более сильных дозах, ведет к большей задержке деления и к гибели клетки при делении. Такое толкование не является, однако, обязательным и, по-видимому, в ряде случаев задержка деления и гибель клеток при возобновлении деления представляют собой независимые явления. Общий метод проверки независимости или связанности этих явлений заключается в том или ином изменении условий опыта, например типа излучения, его интенсивности, температуры, с тем, чтобы установить, носят изменения этих двух эффектов сходный или различный характер. [23]
Для возникновения ранее рассмотренных эффектов радиации ( например, мутаций или разрывов хромосом) необходимо было, чтобы в очень ограниченном районе возникла единичная ионизация или известная концентрация ионизации. В результате эффект всегда сводился к действию одной ионизирующей частицы1, ибо вероятность прохождения нескольких ионизирующих частиц через одно и то же место очень мала. Район, в пределах которого должна-произойти ионизация, необходимая для задержки деления, по-видимому, не столь ограничен, и для ощутимой задержки необходимо несколько ионизации. Поэтому при облучении рентгеновыми лучами этот эффект представляет собой результат действия многих ионизирующих частиц. [24]
Продолжительность задержки зависит от кумулятивной дозы в этот момент. Если облучение произведено несколько раньше, кумулятивная доза к моменту наступления критической стадии успевает снизиться, и, следовательно, эффект оказывается меньшим, чем если бы та же доза была сообщена клетке в момент, более близкий к критической стадии. По данным Карлсона, эта критическая стадия, во время которой эффект данной дозы оказывается максимальным и около которой останавливается митотический процесс при задержке деления клеток, приходится на профазу. Это различие, вероятно, несущественно и лишь отражает различие между разными объектами в отношении легкости, с какой можно отличить раннюю профазу от стадии покоя. Так, на нейробластах кузнечика Карлсон ( 1941Ь) показал, что стадия, описываемая как профаза, продолжается вдвое дольше метафазы, телофазы и анафазы вместе взятых и фактически занимает половину всего промежутка времени между телофазой одного деления и метафазой следующего. В тканях же млекопитающих ( Тэнсли, Спир и Глюксман, 1937) 1 и цыпленка ( Юул и Кемп, 1933; Ласницкий, 1940) продолжительность стадии, описываемой как профаза, равна примерно 1 / 3 продолжительности метафазы, анафазы и телофазы вместе взятых. [25]
Следует иметь в виду, что задержка деления - не единственный эффект, вызываемый в клетке облучением. Если ощутимая задержка деления вызывается дозами, не убивающими клетки, тогда эта задержка может быть изучена. Если же доза, необходимая для того, чтобы вызвать ощутимую задержку, убивает значительную часть клеток, облученных до деления, то изучение задержки деления оказывается невозможным. Нужно, далее, помнить, что термин восстановление употребляется здесь лишь в смысле исчезновения того изменения, которое ответственно за подавление деления, и не подразумевает возвращения клетки к норме во всех отношениях. В ней могут, например, произойти структурные изменения хромосом или другие изменения, которые в конце концов приведут ее к гибели. [26]
У многих объектов облучение вызывает задержку деления, и часть клеток, приступающих к делению после этой задержки, гибнет. Этот факт наводит на мысль, что задержка деления и гибель клеток при делении, возможно, представляют собой по существу сходные явления, различающиеся скорее количественно, чем качественно. Согласно этому предположению умеренное повреждение клетки вызывает некоторую задержку деления или затягивание его ранних стадий, тогда как более сильное повреждение, вызванное облучением в более сильных дозах, ведет к большей задержке деления и к гибели клетки при делении. Такое толкование не является, однако, обязательным и, по-видимому, в ряде случаев задержка деления и гибель клеток при возобновлении деления представляют собой независимые явления. Общий метод проверки независимости или связанности этих явлений заключается в том или ином изменении условий опыта, например типа излучения, его интенсивности, температуры, с тем, чтобы установить, носят изменения этих двух эффектов сходный или различный характер. [27]
Привлекают внимание многочисленные радиобиологические эксперименты с кофеином, являющимся ингибитором фосфодиэстеразы. На Т - клетках почки человека показано, что кофеин ( 1 мМ) при добавлении в среду до и после облучения уменьшает индуцированную радиацией задержку митоза в стадии Ga. Подобным действием обладает МЭА, но при добавлении в среду только до облучения, менее эффективны теофиллин и теобромин. На синхронизированных культурах Physarum polycephalum, облученных в дозе 30 Гр, было продемонстрировано, что модификаторы уровня цАМФ по-разному влияют на задержку деления клеток: кофеин, теобромин и теофиллин уменьшают, дб-ц АМФ, холерный токсин, 1-метил - З - изобутилксантин и параверин не влияют, a RO-20-1724 увеличивает задержку деления. [28]
Привлекают внимание многочисленные радиобиологические эксперименты с кофеином, являющимся ингибитором фосфодиэстеразы. На Т - клетках почки человека показано, что кофеин ( 1 мМ) при добавлении в среду до и после облучения уменьшает индуцированную радиацией задержку митоза в стадии Ga. Подобным действием обладает МЭА, но при добавлении в среду только до облучения, менее эффективны теофиллин и теобромин. На синхронизированных культурах Physarum polycephalum, облученных в дозе 30 Гр, было продемонстрировано, что модификаторы уровня цАМФ по-разному влияют на задержку деления клеток: кофеин, теобромин и теофиллин уменьшают, дб-ц АМФ, холерный токсин, 1-метил - З - изобутилксантин и параверин не влияют, a RO-20-1724 увеличивает задержку деления. [29]
Теория деления - это, пожалуй, самый привлекательный механизм, но и в ней ( как мы увидим) встречаются некоторые трудности. В этой теории постулируются такие условия, что еще на стадии туманности ( или, может быть, позднее) сжимающаяся протозвезда делится на две массы, первоначально образующие контактную систему. Возникшая таким образом система может выглядеть тесной двойной, пока ее компоненты имеют протозвездные размеры, однако ясно, что система неизбежно превратится в широкую двойную, если оба компонента сожмутся до главной последовательности без заметного уменьшения орбитального момента количества движения. Таким образом, и теория деления не способна объяснить происхождение спектрально-двойных. Чтобы объяснить, как образуются тесные двойные, предлагались две возможности: 1) задержка деления на два компонента медленно сжимающейся не достигшей главной последовательности звезды на стадии квазистатического сжатия; 2) постепенная потеря орбитального момента количества движения при сжатии обоих компонентов до главной последовательности. [30]
Страницы: 1 2