Синтез поли
Cтраница 2
Разрывы цепей, возникающие при повреждении ДНК ионизирующей радиацией или вследствие инцизии эндонуклеазами по местам модификации оснований, служат сигналом к синтезу поли - АДФ-рибозы. Связываясь с белками хроматина, полимер модифицирует их структуру, вызывая локальные изменения конденсации хроматина. Эти изменения необязательно включают в себя лишь релаксацию отдельных его участков, повышая тем самым их доступность для нуклеаз инцизии. Возможно и образование сшивок, делающих хроматин более компактным. Такие локаль ные изменения плотности упаковки ДНК в хроматине облегчают координацию отдельных этапов эксцизионной репарации, например, препятствуя чрезмерной активации эндо - и экзонуклаз, не сопровождающейся соответствующей интенсификацией репаративного синтеза. В течение процесса репарации вплоть до полного восстановления двунитевой структуры ДНК поддерживается необходимая интенсивность синтеза поли - АДФ-рибозы, поскольку активность гликогидролазы в присутствии денатурированной ДНК снижена. По окончании репарации ДНК двуни-тевая структура восстанавливается, часть поли - АДФ-рибозы гидро-лизуется гликогидролазои, что сопровождается также восстановлением исходной структуры хроматина. [16]
Фторирование амадиносульфиновой кислоты [478], натрийди-цианамида [479], солей гуанидина [456, 480, 481], азоформамиди-на [482], амидвномочевины [483], бигуавидина [456, 484, 485] и других производных гуанидина [456, 479, 486], а также аммели-на [456, 487] и 3 6-диамино-оош - тетразина [80, 486] в присутствии фторидов натрия, калия или магния широко использовано для синтеза поли ( дифторамино) производных. Хотя селективность указанных реакций невелика, но для некоторых соединений выход удовлетворительный. [17]
Синтез полимеров с возможно более низкой температурой стеклованк имеет важное значение для получения морозостойких каучуков. Синтез поли меров с возможно более высокой температурой стеклования имеет столь ж важное значение для получения твердых теплостойких полимеров, способ ных работать при повышенных температурах и нагрузках. [18]
Показано, что стимуляция синтеза поли - АДФ-рибозы, наблюдаемая при действии повреждающих агентов на ДНК, связана с образованием однонитевых разрывов. Необходимость образования однонитевых разрывов в ДНК для активации синтеза поли - АДФ-рибозы продемонстрирована в модельных экспериментах с минихромосомами вируса SV40 [ Cohen J. J., Berger N. Дальнейшие подтверждения предположения об участии системы АДФ-рибозилирования белков в репарации повреждений ДНК получены в более прямых экспериментах. [19]
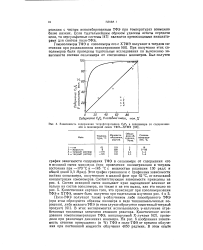 |
Зависимость содержания тетрафторэтилена CsF4 в сополимере от содержания его в мономерной смеси ТФЭ-ХТФЭ. [20] |
ТФЭ при температурах возможно более низких. Если тщательнейшим образом удалены агенты передачи цепи, то персульфатные системы [47] являются превосходными инициаторами для синтеза поли - ТФЭ. [21]
Предложенная схема подтверждена в ряде исследований. Так, возобновлению репликации ДНК в лимфоцитах больных пигментной ксеродермой после УФ-облучения, что происходит после полной репарации димеров тимина, предшествует активация синтеза поли - АДФ-рибозы. [22]
Ферментативный синтез полинуклеотидов может идти и без матрицы. В этом случае полимер получается после латентного периода - продолжительность которого убывает с ростом концентрации фермента. Так идет, например, синтез поли - АТ. Если в качестве затравки-введен оллгомер, то лаг-период быстро убывает с его длиной. Даже короткие олигомеры могут служить матрицами для растущего полимера, который с матрицы соскальзывает. Соответствующая кинетическая теория позволяет получить зависимость времени синтеза полимера от его длины1 и от Скорости отделения цепи от матрицы, па которой она растет. Эта скорость резко убывает при длине оли-гомера, превышающей 4 нук-леотида. [23]
Неизвестным оставался пока порядок расположения нуклеотидов в триплете, так как вероятность всех трех комбинаций УУЦ, УЦУ и ЦУУ одинакова. Но уже изобретены приемы, позволяющие экспериментально изучить и этот вопрос. Принцип одного из них следующий. Для синтеза поли - У берется инициирующая затравка в виде короткого полинуклеотида типа ГГУУ или ААУУ, содержащего дифосфатную группу в 5 - м положении около У. [24]
Предложенная схема подтверждена в ряде исследований. Так, возобновлению репликации ДНК в лимфоцитах больных пигментной ксеродермой после УФ-облучения, что происходит после полной репарации димеров тимина, предшествует активация синтеза поли - АДФ-рибозы. Через 1 - 3 ч после УФ-облучения фибробла-стов человека и периферических лимфоцитов в спейсерных и коро-вых участках хроматина отмечается активация синтеза поли - АДФ-рибозы, состоящей из 15 - 30 звеньев. Это сопровождается активацией репаративного включения метки в ДНК указанных участков. [25]
Поли-А - гомополимер адениловой кислоты, который при наличии подходящих условий синтезируется в различных РНК-полимерных реакциях. Изучение поли - А представляет интерес в связи-с биосинтезом мРНК - Установлено, что мРНК эукариотических клеток оканчивается длинным фрагментом поли - А, который содержит около 200 остатков АМФ, связанных с З - концом полинуклеотидной. Исследования с применением изотопов указывают на то, что этот фрагмент синтезируется в ядре, а присоединяется к мРНК уже после ее транскрипции. В ядрах животных клеток имеются ферменты, катализирующие наращивание З - концацепи мРНК путем присоединения последовательности адениловых нуклеотидов. Известный аналог аденози-на - кордиципин подавляет синтез поли - А и его включение в Д - РНК и мРНК - Он же препятствует накоплению мРН К в полисомах. [26]
Разрывы цепей, возникающие при повреждении ДНК ионизирующей радиацией или вследствие инцизии эндонуклеазами по местам модификации оснований, служат сигналом к синтезу поли - АДФ-рибозы. Связываясь с белками хроматина, полимер модифицирует их структуру, вызывая локальные изменения конденсации хроматина. Эти изменения необязательно включают в себя лишь релаксацию отдельных его участков, повышая тем самым их доступность для нуклеаз инцизии. Возможно и образование сшивок, делающих хроматин более компактным. Такие локаль ные изменения плотности упаковки ДНК в хроматине облегчают координацию отдельных этапов эксцизионной репарации, например, препятствуя чрезмерной активации эндо - и экзонуклаз, не сопровождающейся соответствующей интенсификацией репаративного синтеза. В течение процесса репарации вплоть до полного восстановления двунитевой структуры ДНК поддерживается необходимая интенсивность синтеза поли - АДФ-рибозы, поскольку активность гликогидролазы в присутствии денатурированной ДНК снижена. По окончании репарации ДНК двуни-тевая структура восстанавливается, часть поли - АДФ-рибозы гидро-лизуется гликогидролазои, что сопровождается также восстановлением исходной структуры хроматина. [27]
Вскоре после инфекции вирусный нуклеопротеид оказывается в ядре зараженной клетки, где начинается образование вирусных мРНК - Отметим, что ядерная локализация синтеза вирусных РНК необычна для РНК-содержащих вирусов. Необычен и механизм инициации молекул мРНК - В качестве затравки здесь используются фрагменты транскриптов клеточных генов. Один из вирусных Р - белков узнает кэпированные 5 -концы этих транскриптов и фиксирует их в районе З - концов геномных РНК - Затем этот же или другой Р - белок вносит эндонуклеолитический разрыв в клеточный транскрипт на расстоянии 10 - 13 нуклеотидов от его 5 -конца. Возникающий при этом короткий кэпированный клеточный олиго-нуклеотид и используется как затравка при синтезе () нити. Таким образом, на 5 -концах мРНК вируса гриппа появляется гетерогенный набор олигонуклеотидов клеточного происхождения. При образовании вирусных мРНК ( -) РНК-матрица копируется не полностью. На расстоянии около 20 нуклеотидов от 5 -конца в ней находится олигоуридиловая последовательность длиной 5 - 7 нуклеотидов. После завершения синтеза поли ( А) происходит терминация и около двух десятков 5 -концевых нуклеотидов геномных РНК не представлены в вирус-специфических мРНК - Из образовавшихся таким образом восьми видов вирусных мРНК шесть кодируют по одному белку. Однако часть молекул двух других мРНК подвергается сплайсингу, осуществляемому клеточными ферментами по обычным механизмам. В возникающих в результате сплайсинга дополнительных видах мРНК используются альтернативные открытые рамки считывания, подобно тому, как это характерно для ДНК-содержащих вирусов. [28]
Вскоре после инфекции вирусный нуклеопротеид оказывается в ядре зараженной клетки, где начинается образование вирусных мРНК - Отметим, что ядерная локализация синтеза вирусных РНК необычна для РНК-содержащих вирусов. Необычен и механизм инициации молекул мРНК - В качестве затравки здесь используются фрагменты транскриптов клеточных генов. Один из вирусных Р - белков узнает кэпированные 5 -концы этих транскриптов и фиксирует их в районе З - концов геномных РНК - Затем этот же или другой Р - белок вносит эндонуклеолитический разрыв в клеточный транскрипт на расстоянии 10 - 13 нуклеотидов от его 5 -конца. Возникающий при этом короткий кэпированный клеточный олиго-нуклеотид и используется как затравка при синтезе () нити. Таким образом, на 5 -концах мРНК вируса гриппа появляется гетерогенный набор олигонуклеотидов клеточного происхождения. При образовании вирусных мРНК ( -) РНК-матрица копируется не полностью. На расстоянии около 20 нуклеотидов от 5 -конца в ней находится олигоуридиловая последовательность длиной 5 - 7 нуклеотидов. Как и в случае ВВС, на этом месте РНК-полимераза вируса гриппа буксует и образует З - концевой полиадениловый тракт молекул мРНК - После завершения синтеза поли ( А) происходит терминация и около двух десятков 5 -концевых нуклеотидов геномных РНК не представлены в вирус-специфических мРНК - Из образовавшихся таким образом восьми видов вирусных мРНК шесть кодируют по одному белку. Однако часть молекул двух других мРНК подвергается сплайсингу, осуществляемому клеточными ферментами по обычным механизмам. В возникающих в результате сплайсинга дополнительных видах мРНК используются альтернативные открытые рамки считывания, подобно тому, как это характерно для ДНК-содержащих вирусов. [29]
Страницы: 1 2