Сине-фиолетовая полоса
Cтраница 1
Сине-фиолетовая полоса стоит отдельно и, вероятно, соответствует другому электронному уровню ( табл. 7 и фиг. [1]
Если сине-фиолетовые полосы поглощения экстрактов всех пигментов, содержащихся в диатомовых водорослях, сдвинуть на вышеприведенные величины и графически суммировать, то можно получить составную кривую поглощения, представленную на фиг. Танада не сделал попытки проанализировать область X 620 мр, где все поглощение обусловливается хлорофиллами. Количественное совпадение результатов вычислений с экспериментом не является очень хорошим; вычисленная кривая имеет более высокий пик и более низкую впадину в зеленой области спектра, чем фактическая кривая поглощения клеток. Это может быть обусловлено, по крайней мере частично, рассеянием, хотя кривая поглощения для клеток была получена при помощи спектрофотометра Харди, снабженного интегрирующей сферой, а исследуемые клетки для уменьшения рассеяния были взвешены в глицерине. [2]
Большая интенсивность сине-фиолетовой полосы поглощения ( особенно в хлорофилле Ь) указывает, что естественное время жизни в возбужденном состоянии В несколько меньше, чем в состоянии Y, составляя, вероятно, 5 10 - 8 сек. [3]
Сдвиг в красную сторону сине-фиолетовой полосы хлорофилла а, определенный таким путем, равнялся 8 му. Однако последняя величина была определена по изменению положения наибольшего перегиба кривой поглощения и поэтому не может считаться точной. Положение максимума поглощения фуко-ксантола in vivo было вычислено по разнице между кривыми поглощения клеток до и после экстракции водным метанолом. Оно указывает на сдвиг в красную сторону на 40 му. Положение синего пика хлорофилла с было рассчитано подобным же образом по разнице между спектрами клеток до и после экстракции 50-процентным метанолом; соответствующий сдвиг в красную сторону оказался равным 20 мр. [4]
Мы знаем, однако, что сине-фиолетовая полоса хлорофилла сдвинута приблизительно на тот же интервал, как и красная полоса в шкале частот ( но не длин волн), и что полосы каротиноидов, особенно полосы фукоксантола, вероятно, сдвинуты значительно дальше, чем полосы хлорофилла; поэтому данные фиг. [5]
Эта полоса почти равна по интенсивности сине-фиолетовой полосе, не показанной на фиг. [6]
Сдвиги к красному концу, полученные Танадой для четырех сине-фиолетовых полос поглощения: - 8 Л [ 1 для хлорофилла а и 40 мр для фукоксантола, - можно сравнить с данными более ранних исследований ( см. гл. Данные для хлорофилла а хорошо согласуются между собой. Для фукоксантола сдвиг, найденный Танадой - 40 му. [7]
Характерные изменения, повидимому, имеют место в дублетной структуре сине-фиолетовой полосы. У спиртового раствора хлорофилла а обе компоненты этой полосы хорошо разделены и их пики имеют почти одинаковую высоту, так что, может быть, именно благодаря этому разделению полосы на две почти одинаковые компоненты, сине-фиолетовый пик в этом случае лишь незначительно выше красного. В начале главы уже указывалось, что эти различия свидетельствуют, быть может, о таутомерном равновесии; однако это пока не более, чем предположение. [8]
При наличии стрихнина наблюдаются в стекающей с бихромата калия серной кислоте фиолетовые до сине-фиолетовых полосы. При помощи этой реакции можно обнаружить до 0 001 мг стрихнина; азотная кислота или нитраты мешают определению. [9]
Избирательная сенсибилизация окислительных процессов в фотосинтетическом аппарате, производимая светом, поглощенным каротиноидами или хлорофиллом в сине-фиолетовой полосе, представляет одну из возможностей такого типа. Однако в этом случае подвергаться влиянию должны только те световые кривые, которые принадлежат к типу с оптимумом, зависящим от времени ( этот тип наблюдался, например, у некоторых умбриофильных растений, см. стр. [10]
Если раздражать центр сетчатки красным светом, действующим на fovea centralis retinae, то одновременно в области слепого пятна появляются две сине-фиолетовые полосы, интенсивность которых не изменяется при колебаниях яркости красного света. Франклин считает, что сине-фиолетовые полосы зависят от токов действия сетчатки. Нервные волокна, электрические явления в которых вызывают раздражение соседних чувствительных элементов, находятся в состоянии максимального возбуждения по закону все или ничего. С другой стороны, изменения интенсивности освещения fovea centralis retinae не влияют на электрические явления. Вследствие этого яркость сине-фиолетовых полос остается постоянной, несмотря на изменение силы красного изображения. [11]
Если чашку время от времени наклонять, то в тот момент, когда кислота стекает с кристалликов алкалоида, можно заметить как бы тянущиеся от них сине-фиолетовые полосы. Реакция весьма чувствительна, при помощи ее удается обнаружить 0 001 мг стрихнина. [12]
При судебно-химическом анализе растворяют на крышке от фарфорового тигля выделенное основание стрихнина в концентрированной серной кислоте и добавляют маленький кристаллик дву-хромокислого калия; при покачивании крышечки от кристалла расходятся сине-фиолетовые полосы, постепенно изменяющие окраску через красную до желтой. [13]
Если раздражать центр сетчатки красным светом, действующим на fovea centralis retinae, то одновременно в области слепого пятна появляются две сине-фиолетовые полосы, интенсивность которых не изменяется при колебаниях яркости красного света. Франклин считает, что сине-фиолетовые полосы зависят от токов действия сетчатки. Нервные волокна, электрические явления в которых вызывают раздражение соседних чувствительных элементов, находятся в состоянии максимального возбуждения по закону все или ничего. С другой стороны, изменения интенсивности освещения fovea centralis retinae не влияют на электрические явления. Вследствие этого яркость сине-фиолетовых полос остается постоянной, несмотря на изменение силы красного изображения. [14]
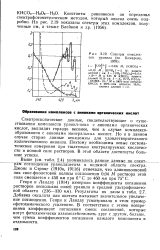 |
Спектры комплексов уранила ( по Комарову, 1959. [15] |
Страницы: 1 2